
Плод винограда в наибольшей степени состоит из какой ткани
Добавил пользователь Дмитрий К. Обновлено: 24.09.2024
Ткань – это совокупность клеток, имеющих общее происхождение, положение и выполняющих общую функцию. Перед тем как разбираться во всем разнообразии тканей высших растений, следует вспомнить строение растительной клетки и ее отличия от животных клеток. Клетки высших растений состоят из клеточной оболочки (клеточной стенки), протопласта (ядра и цитоплазмы) и вакуоли с клеточным соком. В цитоплазме находятся различные органеллы – рибосомы, пластиды, митохондрии, аппарат Гольджи и т.д. Отличительными чертами растительной клетки является наличие целлюлозной клеточной стенки, большой центральной вакуоли с клеточным соком, а также присутствие пластид в цитоплазме. Запасным веществом растительных клеток в отличие от животных является крахмал, а деление клеток происходит с образованием фрагмопласта.
Признаки
Клетки растений
Клетки животных
Немногочисленные крупные с клеточным соком
Многочисленные мелкие пищеварительные или сократительные
С образованием фрагмопласта
Строение растительной клетки. 1 – плазмалемма; 2 – пластида; 3 – клеточная стенка; 4 – цитоплазма; 5 – митохондрия; 6 – плазмодесма; 7 – комплекс Гольджи (диктосомы); 8 — эндоплазматическая сеть; 9 — оболочка ядра; 10 – ядрышко; 11 – ядро; 12 – тонопласт (оболочка вакуоли); 13 – вакуоль.
Ткани высших растений можно классифицировать по-разному. Так, можно различать простые и сложные ткани. Простые ткани сложены одинаковыми клетками. Например, к простым тканям относятся склеренхима, паренхима и хлоренхима. Сложные ткани состоят из разных клеток (проводящих, механических, запасающих). Примерами сложных тканей могут служить флоэма и ксилема. Также ткани можно разделить по происхождению на первичные и вторичные – образовавшиеся в результате деятельности первичных или вторичных меристем соответственно (например, первичная ксилема и вторичная ксилема). Говоря о разнообразии тканей высших растений, чаще всего прибегают к классификации, основанной на их функциях в организме растения. Так, ткани растений разделяют по выполняемым ими функциям на следующие группы:
- покровные (эпидерма, пробка);
- механические (склеренхима, колленхима);
- ассимилирующие (хлоренхима);
- поглощающие (ризодерма, веламен);
- проводящие (ксилема, флоэма);
- запасающие (запасающая паренхима);
- основные (основная паренхима);
- образовательные (апикальная меристема, камбий, феллоген);
- секреторные (железистые волоски, смоляные ходы);
- вентиляционные (аэренхима).
Покровные ткани
Рисунок 1: Эпидерма.
Рисунок 2: Основные типы устьичных аппаратов. 1 – диацитный; 2 –парацитный; 3 –анизоцитный; 4 — аномоцитный.
Вторичная покровная ткань высших растений – это пробка. Пробковый слой обычно образуется на вторично утолщенных стеблях и корнях высших растений. Пробка (она же феллема), образуется в результате работы так называемого пробкового камбия (или феллогена). В феллогене клетки делятся и откладываются наружу, их клеточные стенки утолщаются и суберинизируются (опрбковевают). Суберин – это вещество непроницаемое для воды и воздуха, следовательно, внутреннее содержимое клеток вскоре отмирает. В результате пробковый слой состоит из мертвых клеток и является газо- и водонепроницаемой покровной тканью.
Рисунок 3: Феллема, феллоген, феллодерма.
Механические ткани
Существует две специализированные механические ткани высших растений – склеренхима и колленхима.
Склеренхима, как правило, состоит из клеток вытянутой формы – волокнообразных. Их клеточные стенки утолщаются и лигнифицируются, то есть одревесневают. Живое содержимое клетки впоследствии отмирает. Таким образом, склеренхима – это мертвая ткань, механическую функцию в которой выполняют жесткие клеточные стенки. Склеренхима твердая жесткая ткань и в растении она выполняет армирующую функцию, располагаясь обычно тяжами или слоями. Однако иногда склеренхима может быть представлена в виде отдельных клеток с одревесневшими клеточными стенками, разбросанных в толще некой мягкой ткани (например, паренхимы). Такие клетки называются склереидами. По форме различают разные типы склереид: брахисклереиды, астросклереиды, остеосклереиды и волокнистые склереиды. Все склеренхимные элементы вместе составляют стереом – совокупность всех толстостенных одревесневших клеток растения. Следует также помнить, что отчасти механическую функцию, подобно склеренхиме, выполняет водопроводящая ткань ксилема (в особенности ядровая древесина – вторичная ксилема, прекратившая проводить воду).
Рисунок 1: Склеренхима.
Рисунок 2: Колленхима. А – рыхлая; Б – пластинчатая; В – уголковая. 1 – первичная; клеточная стенка; 2 – вторичная клеточная стенка; 3 – межклетник; 4 – протопласт.
Ассимилирующие ткани (хлоренхима)
Рисунок 1: Хлоренхима.
Рисунок 2: Поперечный срез листа. 1 – эпидерма; 2 – столбчатая хлоренхима; 3 – губчатая хлоренхима; 4 – подустьичная полость; 5 – устьице.
Поглощающие ткани
Высшие растения поглощают воду с помощью специальных тканей. У мохообразных отсутствуют корни, и всасывание воды происходит всей поверхностью тела (например, с помощью гиалиновых клеток у сфагновых мхов) или с помощью ризоидов – длинных тонкостенных клеток. Сосудистые растения имеют корни, поверхность которых покрыта ризодермой (эпиблемой) – специализированной всасывающей тканью. Ризодерма гомологична эпидерме, то есть также формируется из одного внешнего слоя клеток, покрывающих орган. Однако ризодерма не является покровной тканью, поскольку практически не выполняет защитную функцию. Ее клетки тонкостенные и специализируются на поглощении воды и минеральных солей из почвы, поглощение при этом происходит избирательно и с затратой энергии. В ризодерме различают два типа клеток: трихобласты и атрихобласты. У трихобластов наружная часть клетки выпячивается и образует длинный вырост – корневой волосок, служащий для увеличения поверхности всасывания. Корневой волосок выделяет слизь, которая помогает растворять поглощать минеральные вещества из почвы. Атрихобласты не формируют корневых волосков, но также поглощают вещества своей поверхностью.
Рисунок: Ризодерма. А – Продольный разрез корня; Б – Клетки ризодермы. 1 – зона проведения; 2 – зона всасывания; 3 – зона роста; 4 – зона деления; 5 – корневые волоски; 6 – корневой чехлик.
У некоторых тропических эпифитных растений вместо ризодермы развивается веламен. Веламен гомологичен ризодерме, но в отличие от нее является многослойной тканью и состоит из отмерших клеток. Их клеточные стенки имеют спиральные утолщения, которые служат ребрами жесткости, сами клеточные стенки частично разрушаются, а внутреннее содержимое клеток отмирает. В результате получается структура наподобие губки, которая способна впитывать воду из влажного воздуха, тумана или осадков. Таким образом, веламен поглощает вещества пассивно и не избирательно. Направленный и избирательный транспорт воды дальше внутрь корня происходит при участии экзодермы, подстилающей веламен (как, впрочем, и любую ризодерму).
Проводящие ткани (ксилема, флоэма)
Ксилема – сложная ткань, то есть состоит из клеток разной морфологии. В состав ксилемы одновременно входят и проводящие, и механические, и запасающие элементы.
Ксилема проводит воду с растворенными в ней минеральными веществами от корней по всему остальному телу растения. Таким образом, по ксилеме в основном осуществляется восходящий ток. Проводящие элементы ксилемы – это сосуды и трахеиды. Следует помнить, что ксилема голосеменных растений лишена сосудов. Трахеида образуется из клетки удлиненной формы, ее клеточная стенка утолщается и лигнифицируется, то есть одревесневает. Протопласт при этом отмирает и в результате получается мелкий капилляр, по которому может транспортироваться вода. Прочные клеточные стенки предохраняют просвет капилляра от схлопывания. От трахеиды к трахеиде вода транспортируется через специальные поры. Сосуд, по сути, является таким же капилляром, как и трахеида, но более длинным, широкопросветным и многоклеточным. Каждый сосуд состоит из отдельных клеток (члеников сосуда) с одревесневшей оболочкой и отмершим протопластом, между члениками сосуда формируются уже не поры, а перфорационные пластинки (то есть сквозные отверстия). Между сосудами, как и между трахеидами, есть поры, через которые также может транспортироваться вода. Кроме проводящих элементов, в состав ксилемы входят механические волокна – волокна либриформа. Это удлиненные клетки, похожие на трахеиды, однако их клеточные стенки очень сильно утолщены и лигнифицированы. Просвет таких капилляров слишком мал для осуществления транспорта воды, зато толстая и прочная клеточная стенка выполняет механическую функцию подобно склеренхиме. Ксилема в основном состоит из мертвых клеток, обычно небольшой процент живых клеток представлен древесинной паренхимой. Эти клетки в основном выполняют запасающую функцию.
Флоэма, как и ксилема, – это сложная ткань, которая состоит из разных клеток. В состав флоэмы входят проводящие механические и паренхимные (в том числе запасающие) элементы.
Рисунок: Проводящие ткани. А – ксилема; Б – флоэма. 1 – сосуды ксилемы; 2 – трахеиды; 3 – клетки древесной паренхимы; 4 – поры; 5 - ситовидные трубки; 6 – клетки – спутницы; 7 – ситовидные поля; 8 – клетки лубяной паренхимы.
Запасающие ткани (запасающая паренхима)
Запасающие ткани высших растений бывают различными по происхождению, также различия заключаются в том, какие именно вещества и в какой части клетки запасаются.
Главное запасное вещество высших растений – это крахмал. Крахмал синтезируется и откладывается в виде зерен в специальных пластидах – амилопластах. Крахмальные зерна увеличиваются в размере и растягивают пластиду. В результате клетка такой запасающей ткани содержит множество крупных зерен крахмала – примером может служить запасающая ткань в клубне картофеля.
Если растение запасает питательные вещества не на очень долгий срок, то они могу откладываться в виде сахаров в вакуолях клеток. Например, в сочной ткани многих плодов. Сочный плод рассчитан на то, что его съест некое животное, а значит, он должен быть привлекательным для него – питательным и сладким.
В эндосперме некоторых семян запасание происходит за счет утолщения клеточной стенки, в которой откладывается гемицеллюлоза.
При прорастании семени клетки частично растворяют свои клеточные стенки и потребляют углеводы, из которых она состоит. В качестве запасного вещества может выступать белок. Он может откладываться в вакуолях (алейрон) или в лейкопластах. В цитоплазме запасаются жиры в виде сферосом.
Кроме питательных веществ, ткань может запасать воду. Клетки водоносной ткани бывают ослизнены и имеют крупные вакуоли, в которых сохраняется влага.
Рисунок: Запасающая паренхима клубня картофеля. 1 – крахмальные зерна.
Основные ткани (основная паренхима)
К системе тканей основной паренхимы традиционно относят все ткани, образованные из основной меристемы (не являющиеся покровными и проводящими) то есть запасающие, фотосинтезирующие и т.д. Однако эти ткани специализированы на выполнении конкретной функции и рассматриваются обычно отдельно. Основной паренхимой в узком смысле называют ткань, состоящую из рыхло расположенных более или менее шарообразных клеток.
Между клетками есть заметные межклетники.
Данная ткань не специализирована для выполнения какой-то определенной функции, это структурная ткань, заполняющая пространство того или иного органа. Поскольку клетки основной паренхимы живые, их клеточные стенки не лигнифицированы, а в цитоплазме есть полный набор клеточных органелл, при необходимости она может становиться запасающей, водоносной или фотосинтезирующей тканью.
Также основная паренхима может проявлять меристематическую активность – клетки могу начать делиться. Со временем клеточные стенки паренхимы могут одревесневать, тем самым начиная выполнять механическую функцию.
Таким образом, основная паренхима – это неспециализированная структурная ткань, которая может специализироваться при определенных условиях.
Рисунок: Основная паренхима.
Образовательные ткани
Массив ткани, в которой происходят клеточные деления в теле высшего растения, следует назвать образовательной тканью или меристемой. Образовательные ткани не являются постоянными. Клетки меристемы недифференцированные и не специализированные, у них тонкие клеточные оболочки. Данные клетки делятся и в дальнейшем преобразуются в ту или иную специализированную ткань.
Высшие растения имеют верхушечный рост, их побеги (и корни) нарастают за счет верхушечной или апикальной меристемы. Рассмотрим апикальную меристему стебля. Это массив делящихся клеток на вершине растущей оси побега, ниже апекса ткань разделяется на три отдельные меристемы: протодерму, прокамбий и основную меристему. Протодерма – это один поверхностный слой клеток меристемы. Из протодермы в дальнейшем формируется эпидерма. Прокамбий представлен тяжами клеток, которые дифференцируясь, становятся проводящими тканями (формируют проводящие пучки). Остальные ткани стебля (паренхима, хлоренхима, склеренхима и т.д.) формируются из основной меристемы.
Рисунок 1: Апикальная меристема стебля.
У высших растений выделяют две вторичные латеральные меристематические ткани – камбий и феллоген. Камбий (или сосудистый камбий) закладывается в проводящих пучках стебля или корня между флоэмой и ксилемой. В результате клеточных делений внутрь откладывается ткань, дифференцирующаяся в ксилему, а наружу – будущая флоэма. За счет работы камбия происходит процесс вторичного утолщения стебля или корня. Соответственно, сформированные камбием проводящие ткани будут называться вторичными – вторичная ксилема и вторичная флоэма. Следует помнить, что при вторичном утолщении камбиальная зона возникает не только внутри проводящих пучков, но и формируется так называемый межпучковый камбий. В результате на поперечном срезе камбий имеет вид общего меристематического кольца.
Рисунок 2: Камбий. 1 – эпидерма; 2 – паренхима; 3 – флоэмные волокна; 4 – флоэма; 5 – пучковый камбий; 6 – ксилема; 7 – межпучковый камбий.
Феллоген (или пробковый камбий) возникает в корнях и стеблях растений при их вторичном утолщении. При утолщении эпидерма и впоследствии первичная кора опадает и отмирает, покровную функцию в данном случае выполняет пробковый слой, формируемый феллогеном. В результате клеточных делений в феллогене, наружу откладываются клетки феллемы (или пробка). Феллоген снизу подстилается слоем клеток – феллодермой. Комплекс из трех данных тканей носит название перидерма.
Рисунок 3: Феллема, феллоген, феллодерма.
Секреторные ткани (железистые волоски, смоляные ходы)
Секреторные (или выделительные) структуры высших растений очень разнообразны как по строению, так и по происхождению. Они делятся на две группы: экзогенные и эндогенные.
Экзогенные секреторные структуры расположены на поверхности тела растения. К ним относятся гидатоды – структуры, выделяющие капельно-жидкую воду. Их наличие характерно для растений, обитающих в условиях повышенной влажности. К гидатоде подходят проводящие элементы ксилемы, по которым транспортируется вода. Также к экзогенным структурам относятся различные железистые волоски или более крупные многоклеточные железки. Они, как правило, выделяют эфирные масла, которые скапливаются под кутикулой наружных клеток структуры. Нектарники также являются экзогенными секреторными структурами. Они выделяют секрет богатый сахарами, сахара поступают в нектарники по флоэмным элементам. Различают флоральные (расположенные в цветке) и экстрафлоральные нектарники.
Эндогенные секреторные структуры находятся внутри тела растения. Они бывают одноклеточные и многоклеточные. Одноклеточные структуры могут быть разнообразными по содержанию – это слизевые, кристаллоносные, масляные клетки, одноклеточные млечники, а также прочие клетки, накапливающие в себе те или иные вещества. Многоклеточные эндогенные структуры обычно выделяют секрет в некую полость, представляющую собой межклетник. По типу межклетников различают схизогенные и лизигенные вместилища. По типу содержащегося в них секрета различают смоляные, слизевые, камеденосные ходы и т.д. К многоклеточным структурам также относят млечники. Они состоят из трубчатых клеток, внутри которых находится млечный сок. Если концевые стенки трубчатых клеток деградируют, то такой млечник называют нечленистым.
Рисунок: Секреторные структуры.
Вентиляционные ткани (аэренхима)
Аэренхима – это вентиляционная ткань или ткань проветривания. Главную функцию аэренхимы выполняют крупные межклетники, по которым и циркулирует воздух. Воздух необходим высшим растениям как для дыхания, так и для процессов фотосинтеза. Наличие аэренхимы характерно для водных или околоводных высших растений. Воздух, находящийся в системе полостей аэренхимы, не только вентилирует все части растения (в особенности подводные), но и придает им плавучесть, как, например, листьям кувшинки.
Рисунок: Аэренхима. А – аэренхима на поперечном срезе стебля; Б – клетки пленчатой перегородки, разделяющей полости аэренхимы; В – аэренхима из трехмерно расположенных звездчатых клеток.
Вентиляционная ткань выполняет свою функцию за счет многочисленных увеличенных межклетников. Стоит помнить, что межклетники по типу происхождения делятся на три типа. Схизогенные межклетники образовались в результате простого расхождения клеток в пространстве. Лизигенные полости формируются в результате деградации (лизиса) некоторых клеток. Крупные рексигенные полости являются результатом механического разрыва тканей, например, в центре черешков или стеблей некоторых растений.
Различные типы покровных тканей выполняют в некоторой мере и механические функции, защищая нежные ткани, расположенные ковнутри органа.
Но есть еще специальные клетки и слагаемые из них ткани, которые выполняют механические функции. Такие клетки и ткани называют механическими. Эти клетки и ткани усиливают противодействие всего растения или его органов, прежде всего излому или разрыву.
Механические клетки и ткани имеются во всех органах каждого растения, произрастающего в наземных условиях. В сильно растущих в длину частях растения, например в стеблях, элементы механических тканей также вытянуты, имеют прозенхимный характер. В органах со слабо выраженным ростом в длину механические клеточные элементы более или менее изодиаметричны — паренхимного характера. Такие элементы встречаются в плодах и листьях. Разумеется, абсолютно строгого разграничения между типами механических элементов не существует: нередко в стеблях имеются механические паренхимные клетки, а в листьях — вытянутые прозенхимные.
Все механические ткани по ряду признаков могут быть разделены на несколько групп, из которых каждая отличается определенными характерными чертами. Таких основных групп три: колленхима, лубяные и древесинные волокна и склереиды.
Колленхима встречается в стеблях, а также нередко в черешках и средних жилках листьев, реже в цветоножках и в плодоножках. Она находится в тех частях органов, где расположены сочные ткани, состоящие из живых клеток, и преимущественно свойственна двудольным растениям; у однодольных она встречается реже.
Колленхима служит для укрепления растущих органов. Она состоит из живых клеток, содержащих хлоропласты. Обычно колленхимные клетки длинные, иногда достигают значительной длины (2 мм), но среди них есть и вполне паренхимные клетки. Своеобразны утолщения оболочек колленхимных клеток. Наиболее часто наблюдается так называемое уголковое утолщение, когда полосы утолщений тянутся по углам клеток. Если утолщены тангентальные стенки клеток, то образуется пластинчатая колленхима. Характер утолщений колленхимы в значительной степени зависит от самого расположения клеток. Если клетки колленхимы расположены радиальными рядами, то утолщения возникают на тангентальных стенках. При беспорядочном расположении клеток утолщения концентрируются по углам.
Оболочки колленхимных клеток целлюлозные и на поперечных разрезах органов выделяются особым блеском. Особенность оболочек колленхимных клеток состоит в том, что в них содержится большое количество воды. Возможно, что это благоприятствует вытягиванию оболочки при росте органа.
Колленхима расположена почти всегда по периферии органа. Она дифференцируется раньше других механических тканей. Располагается колленхима или сплошным слоем в несколько рядов клеток вблизи периферии органа или собрана отдельными пучками, связанными с пучками проводящей системы. Если на стеблях есть ребра, то колленхима в виде компактных пучков тянется вдоль стебля по ребрам (например, зонтичные).
Колленхима принадлежит к числу простых тканей, т. е. клетки ее не смешиваются с клетками другого характера. Поэтому границы колленхимы обычно очерчены довольно резко. Однако иногда образуются переходные формы от колленхимных клеток к клеткам паренхимы.
Клетки колленхимы соприкасаются друг с другом тупо заканчивающимися концами и имеют простые поры. Клеточные полости их всегда широкие. Благодаря хлоропластам, колленхимные клетки, подобно соседним паренхимным клеткам коры, могут участвовать в фотосинтетической деятельности, но в значительно меньшей степени.
Лубяные волокна представляют собой вытянутые по оси органа длинные толстостенные клетки. Длина их у разных видов сильно колеблется. У конопли лубяные волокна имеют в среднем длину около 10 мм, у льна — около 40 мм, у крапивы — около 80 мм. Самые длинные волокна были найдены у субтропического прядильного растения рами — до 500 мм. У специальных текстильных сортов льна найдены лубяные волокна до 120 мм длины. У некоторых растений оболочки лубяных волокон остаются целлюлозными, но в большинстве случаев они более или менее рано одревесневают. Однако иногда одревеснение волокон происходит лишь у основания стебля.
К растениям с целлюлозными оболочками лубяных волокон принадлежат лен, рами. У конопли же, особенно у некоторых сортов ее, а также у крапивы оболочки лубяных волокон скоро одревесневают на всем протяжении стебля.
В молодом состоянии, во время роста, лубяные волокна содержат в своих полостях жизнедеятельный протопласт со многими ядрами. После окончательного формирования лубяного волокна протопласт обычно отмирает, остатки его лишь кое-где рассеяны в клеточной полости.
В живом протопласте лубяных волокон иногда обнаруживаются крахмальные зерна, но клеточная полость их обычно очень узка. Только у молодых волокон клеточная полость выражена отчетливо, у старых же, лишенных протопласта, полость почти совершенно исчезает. В толстых стенках иногда довольно хорошо заметна слоистость. Толщина оболочки даже у вполне сформировавшихся волокон не по всей длине одинакова: утолщенные участки перемежаются с утонченными, поэтому полость волокна местами суживается и расширяется.
У некоторых растений (виноградная лоза) полость лубяных волокон перегорожена тонкими перегородками. Такие анатомические элементы называются перегородчатыми лубяными волокнами.
Весьма характерно смыкание волокнистых клеток между собой.
Каждое волокно своими острыми концами вклинивается между другими волокнами. Такое взаимное соединение волокон, отличающееся от взаимного соединения паренхимных клеток, способствует наибольшей прочности ткани и происходит вследствие так называемого скользящего роста концов таких клеток.
На оболочках лубяных волокон имеются простые поры, расположенные по спирали. Последнее указывает на спиральную текстуру микрофибрилл во вторичной оболочке волокна. Спиральное расположение фибрилл придает волокну особую прочность. На спиральность текстуры указывает также то, что при механическом разрушении оболочки (сильное надавливание, приготовление срезов, поражение микроорганизмами) концы поврежденных оболочек иногда развертываются в виде вытянутой спирали.
Обычно лубяные волокна составляют простую ткань, т. е. к ним редко примешиваются другие анатомические элементы. Расположены лубяные волокна либо более или менее широким поясом, как в стеблях некоторых растений, либо собраны отдельными группами, образующими вместе с проводящими тканями сосудисто-волокнистые пучки.
Но есть и такие растения, у которых лубяные волокна не составляют отдельных пучков или компактных массивов, а перемешаны с паренхимными клетками. Такая паренхима называется лубяной.
При кажущейся простоте строения лубяные волокна по своей организации сильно варьируют даже у одного и того же растения. В нижней части стебля пучки лубяных волокон менее обособлены, чем в верхней части. Длина лубяных волокон в различных местах стебля одного и того же растения неодинакова: в нижней части расположены более короткие волокна, чем в средней и верхней частях стебля.
При произрастании растений одного и того же вида в различных климатических условиях создаются так называемые географические расы, своеобразные особенности которых отчасти передаются по наследству. У конопли, как и у других культурных растений, известно много географических рас. Расы конопли отличаются как количеством волокна (степенью волокнистости), так и качеством его. Анатомическое исследование обнаруживает значительную разницу и в строении лубяных волокон у географических рас конопли различного происхождения.
Лубяные волокна, подобные только что рассмотренным волокнам конопли, образуются перициклом — образовательной тканью, расположенной по периферии осевого органа, на границе с первичной корой. Лубяные волокна, возникшие в перицикле, называют первичными. Есть и вторичные лубяные волокна. Они возникают в результате деятельности другой образовательной ткани — камбия. В стеблях конопли, кроме первичных лубяных волокон, образуются и вторичные. То же наблюдается у кенафа, канатника, кендыря, рами — южных текстильных лубо-волокнистых растений. У льна и подсолнечника лубяные волокна только первичные.
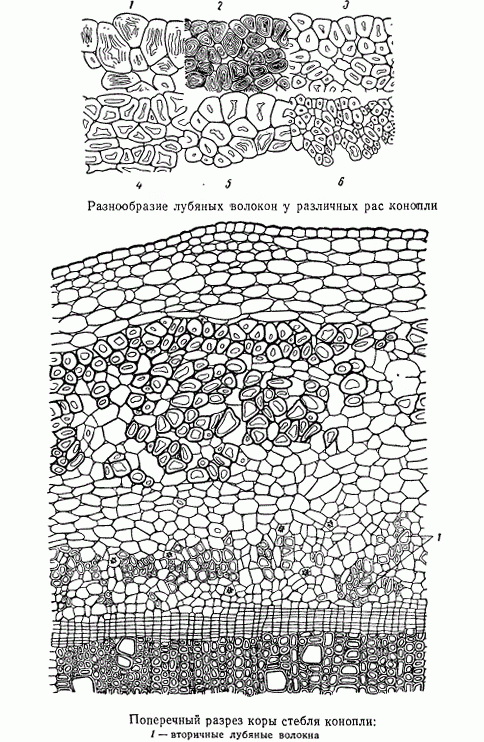
Вторичные лубяные волокна травянистых растений обычно развиты значительно слабее первичных. У конопли, например, вторичные лубяные волокна, по сравнению с первичными, имеют более мелкий просвет и одревесневшие оболочки, оболочки же первичных лубяных волокон многих сортов конопли остаются неодревесневшими или одревесневают весьма слабо. Текстильное значение имеют только первичные волокна и не только у конопли, но и у других, произрастающих в умеренном климатическом поясе травянистых текстильных растений. Вторичные лубяные волокна у этих растений наиболее обильно образуются лишь в базальной части стебля. Чем выше вдоль по стеблю, тем меньше развивается вторичных лубяных волокон; примерно на половине длины стебля они уже не образуются.
У древесных растений, наоборот, наиболее обильно развиты вторичные лубяные волокна. Первичные лубяные волокна, если и образуются, то лишь на ранней стадии развития, когда еще функционирует перицикл. Вторичные лубяные волокна в отличие от первичных, по-видимому, у всех древесных растений имеют одревесневшие оболочки. Примером вторичных лубяных волокон могут служить волокна липы, составляющие прочную основу так называемого лыка. Вторичные лубяные волокна никогда не достигают такой длины, как первичные.
Прочность лубяных волокон, особенно первичных, общеизвестна. Недаром из них выделывают различные ткани, канаты и пр. Волокна некоторых растений не уступают по прочности стали, далеко оставляя ее за собой по эластичности. Разумеется, колебания в прочности среди лубяных волокон тоже значительны и зависят от многих условий (степени одревеснения оболочек, влажности и пр.).
Больше всего лубяных волокон развивается в стеблях, в корнях их значительно меньше, часто не бывает совсем. У некоторых растений, преимущественно у однодольных (юкки, драцены, некоторые пальмы и пр.), хорошо развитые лубяные волокна встречаются в листьях. Есть лубяные волокна и в плодах различных типов (бобах, коробочках и др.).
Волокна, находящиеся в древесной части стебля, называются либриформом. Если лубяные волокна представляют собой самые толстостенные элементы коры, то ткань либриформа состоит из самых толстостенных анатомических элементов древесины. В основе своего строения либриформ сходен с лубяными волокнами и назначение его, по-видимому, такое же. Волокна либриформа значительно короче лубяных волокон (не больше 2 мм). Оболочки клеток либриформа всегда одревесневшие, снабжены простыми порами со щелевидными очертаниями. Так же, как и у лубяных волокон, имеющих простые поры, поры либриформа расположены по спирали. У некоторых растений (виноград) полость волокон либриформа, подобно полости лубяных волокон, перегорожена тонкими перегородками, не разделяющими, однако, окончательно самой клетки. Такие волокна либриформа, так же как и лубяные, называют перегородчатыми.
По своей структуре и внешнему виду клетки либриформа иногда представляют собой переход к другим типам тканей. Они могут быть очень короткими, приближаясь по форме к паренхиме. Оболочки таких клеток имеют простые поры, их протопласт долго остается живым, в них периодически накопляется крахмал. Но есть клетки либриформа, приближающиеся к водопроводящим элементам — трахеидам и сосудам. Оболочки таких клеток имеют окаймленные поры, протопласт в них скоро разрушается. Вообще тип либриформа не столь отчетлив, как тип лубяных волокон.
Либриформ также весьма распространен среди высших растений, как и лубяные волокна (твердый луб). У лиственных деревьев либриформ иногда занимает значительную часть древесины, в особенности в тех массивах ее, которые образовались во вторую половину вегетационного периода.
На примере либриформа особенно отчетливо выявляется общераспространенный принцип, обнаруживаемый при рассмотрении почти каждого анатомического элемента в растении, — принцип разнообразия функций, выполняемых той или другой клеткой или тканью. Либриформ наряду с основной функцией выполняет еще функции хранилища запасов и проведения воды. В соответствии с этим внешний вид и самая структура клеток либриформа не вполне устойчивы: в ряде случаев есть немало переходных форм, имеющих черты, характерные уже для других анатомических элементов.
Склереиды представляют собой клетки самой разнообразной формы, толстостенные, одревесневшие, не слишком большой длины или чаще паренхимные. Они встречаются в различных органах растений: плодах, листьях, стеблях. Наиболее типичным образцом склереид могут служить каменистые клетки, представляющие собой пример опорных элементов. В плодах груши и айвы каменистые клетки выполняют роль опоры для сочной мякоти зрелого плода. Каменистые клетки обычно паренхимные. Более вытянутые элементы встречаются в плодоножках. Твердые оболочки различных орехов, пергаментный слой в бобах также состоят из склереид. Нередко группы таких клеток способствуют раскрыванию плодов, разрыву более тонких стенок.
Иногда вытянутые склереиды ветвятся. Ветвистые склереиды чаще всего размещены одиночно и относительно велики. Неветвистые склереиды собраны группами, образуя подобие простой ткани. Нечто сходное происходит и с либриформом. Чаще всего клетки либриформа собраны группами, но нередко маленькие группы клеток либриформа или даже отдельные клетки перемежаются с клетками другого характера.
Такие ткани, состоящие из разнообразных анатомических элементов, можно назвать смешанными.
Механические ткани в каждом органе сконцентрированы таким образом, что обусловливают прочность на разрыв или изгиб в определенном направлении. В стебле, подвергающемся преимущественно изгибу, механические ткани расположены по периферии, что особенно ясно выражено у травянистых растений.
В корнях, подвергающихся преимущественно растяжению, наоборот, периферическая зона (кора) почти лишена механических элементов, и они расположены ближе к центру оси корня.
На основе анализа закономерностей в распределении механических тканей в растениях некоторые исследователи разработали ряд схем конструкций органов растений с использованием строительно-механических принципов. Следует, однако, признать, что строительно-механический принцип объясняет лишь одну сторону функционирования системы механических тканей.
Дело в том, что механические анатомические элементы редко выполняют только механические функции. Только с этих позиций трудно объяснить, например, значение незначительного по толщине кольца лубяных волокон в древесном стволе с его солидной древесиной и притом нередко весьма прочной.
Структура и функции каждого органа сложны и разнообразны, в силу чего с помощью чисто механических схем нельзя выяснить закономерности в организации структуры растений, тем более что между различными анатомическими элементами существует ряд промежуточных форм. Структура пластична. В живом организме структура — процесс.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Виноградная гроздь. Представляет собой соплодие, состоящее из механически прочного остова — гребня и нежных плодов — ягод, прикрепленных к гребню. Формируется гроздь из соцветия в процессе его роста и развития после цветения. При этом ножка соцветия преобразуется в ножку грозди, ось соцветия с разветвлениями — в гребень, завязи — в ягоды.
Ягода винограда. Это — сочный нераскрывшийся плод виноградного растения, который служит для защиты и распространения заключенных в ней семян. Ягода состоит из тонкой эластичной кожицы, охватывающей разросшийся околоплодник — мякоть. Мякоть состоит из крупных клеток с большими вакуолями, наполненными клеточным соком.
Ягода в разных направлениях пронизана сосудисто-волокнистыми пучками, которые на периферии мякоти образуют мелкую сеть, хорошо видимую, если снять кожицу. Часть проводящих сосудистых пучков связаны с центром ягоды. По ним из плодоножки в ягоду поступают питательные вещества.
Механическую прочность прикрепления ягоды к плодоножке обеспечивает утолщение растительной ткани — подушечка. Семена ягоды заключены в оболочку мякоти, состоящую из мелких клеток, плотно примыкающих друг к другу.
Кожица зрелой ягоды покрыта восковым прюиновым налетом. В прилегающих к кожице слоях эпидермиса находятся различные красящие вещества, определяющие окраску ягод. У сортов, имеющих окрашенный сок, антоцианы, кроме того, распространены по всей мякоти. Окраска ягод имеет исключительное разнообразие оттенков: от желто-зеленой (белой), серой и розовой до красной и темно-синей (черной), что обусловлено цветовой гаммой красящих веществ.
В клетках ягоды сахара, кислоты, эфирные масла и другие ценные вещества распределяются неравномерно. Эфирные масла, к примеру, накапливаясь по всей мякоти в особых вместилищах, концентрируются ближе к кожице. Сахара, наоборот, в наибольшем количестве находятся в рыхлых сочных клетках мякоти и составляют основу сусла-самотека. В перезрелом же винограде сок становится настолько вязким и густым, что прессовые фракции сусла оказываются наиболее сладкими.
Существует корреляция между числом семян в ягоде и содержанием в ней сахаров и кислот: чем больше в ягоде семян, тем меньше в ней сахаров и выше кислотность сока. Бессемянные ягоды кишмишных сортов винограда — самые сладкие и малокислотные.
Еще совсем недавно Ксения Жучкова жила и работала в нашем городе. Девушка закончила факультет Экономики и управления УПИ, трудилась в страховой и энергетической компаниях, на металлургическом заводе и в Почетном Консульстве Италии в Екатеринбурге.
С позволения автора мы будем публиковать некоторые тексты Ксении в виде познавательных уроков. Уверены, эти материалы придутся по вкусу всем, кто любит вино и интересуется им. Обратите внимание, в конце каждого текста будет небольшой толковый словарь, в котором вы сможете найти все незнакомые слова.

На данный момент учеба практически осталась позади, так что у меня появилось время осуществить задуманное. В своих текстах я хочу вспомнить то малое, что удалось услышать на лекциях и самостоятельно углубиться в самое интересное для меня - создание вина.

Наша группа в Университете была разношерстная. За одной партой с дипломированными энологами сидели примерно такие, как я - экономисты, философы и финансисты, поэтому мы изучали даже самые элементарные вещи.
Начнем, пожалуй, с винограда. Из чего он состоит? Из гребня, который занимает от 3 до 7% веса грозди, косточек (составляют от 0 до 6% веса ягоды), кожицы (составляет от 8 до 20% веса ягоды) и мякоти (составляет от 75 до 85 % ягоды).
А теперь обо всем по порядку.

Виноградные косточки
Косточки являются важным источником фенолов*, участвующих в винификации* красных вин. В косточках, в зависимости от сорта винограда, содержится от 20 до 55% всех фенолов ягоды.
Во время созревания винограда количество танинов* в косточке уменьшается и увеличивается их уровень полимеризации, что понижает вязкость. Грубо говоря, чем более спелый виноград, тем менее вязкой будет на вкус косточка, которую мы раскусим. Именно поэтому в виноделии очень важно определять степень зрелости косточек, чтобы собрать виноград в правильный момент.
.jpg)
Виноградная кожица
Кожица, в основном, нужна для аромата и фенолов, количество которых увеличивается при созревании ягоды. Фенолы в данном случае – это бензойные и коричнокислые кислоты, флавоноиды* и танины, а также антоцианы* в кожице красного винограда.
Большинство ароматов содержатся именно в кожице. На профессиональном языке ее называют экзокарпием*. Ароматы высвобождаются в момент разрывания кожицы и претерпевают различные трансформации в процессе винификации.
Кроме того, именно на кожице винограда находятся дрожжи и бактерии, без которых невозможен сам процесс винификации.

Мякоть винограда
Мякоть представляет самую большую часть ягоды - от 75 до 85%. Вакуолярное содержимое клеток мякоти представляет собой не что иное, как сусло.
Основные составляющие мякоти: сахара (глюкоза и фруктоза), органические кислоты (винная, лимонная и яблочная), минеральные соли (калий, кальций, магний, натрий) и азот.

Аромат вина
Все сорта винограда делятся на три группы, исходя из количества монотерпенов, которые они содержат. Бывает мускатный виноград (гевюрцтраминер, различные мускаты), немускатный ароматный виноград (траминер, кернер, рислинг и т.д.) и нейтральные сорта, ароматность которых не зависит от количества содержащихся в них монотерпенов (каберне совиньон, шардоне, мерло, совиньон блан, сира, треббьяно и т.д.).
.JPG)
Созревание винограда
Процесс созревании винограда - это период от момента перемены виноградом цвета до момента его сбора. Очень многое меняется за это время. Например, количество сахара увеличивается, а кислотность, в основном, понижается. По соотношению между количеством сахара и общей кислотностью можно получить представление о степени зрелости винограда и принять решение о времени его сбора.
Но кислотность и содержание сахара не единственные важные параметры, которые нужно держать под контролем. Важными являются также ароматность, то есть количество монотерпенов и содержание фенолов.

Толковый словарь:
Фенолы - органические соединения ароматического ряда, в молекулах которых гидроксильные группы связаны с атомами углерода ароматического кольца. Фенолы в вине могут реагировать с кислородом и другими веществами, что и приводит к развитию сложного аромата и цвета вина.
Винификация - процесс превращения винограда в вино, при котором естественное брожение происходит под наблюдением и контролем специалиста, целью которого является создание конкретного конечного продукта.
Флавоноиды - это крупнейший класс растительных полифенолов. Флавоноиды известны как растительные пигменты.
Антоцианы – пигментные вещества из группы гликозидов. Они находятся в растениях, обусловливая красную, фиолетовую и синюю окраски плодов и листьев.
Экзокарпий - внеплодник, наружный слой околоплодника растений.
Терпены - ненасыщенные углеводороды, молекулы которых построены из изопреновых звеньев. В большом количестве терпеновые соединения встречаются в винограде, сусле и вине.
Читайте также: