
У покрытосеменных синергиды это
Добавил пользователь Алексей Ф. Обновлено: 19.09.2024
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Навашиным в 1910 г. на лилейных растениях.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит — это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Схема мегаспорогенеза (1-5) и мегагаметогенеза (6-12) у растений
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры. В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других — в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения. Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно. В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других — из двух (биспорический тип) и четырех спор (тетраспорический тип).
Типы развития зародышевых мешков
Как мы указывали, при моноспорическом типе развивается лишь одна мегаспора из четырех, а остальные три разрушаются подобно тому, что имеет место с редукционными тельцами у животных. При других типах развития зародышевого мешка сохраняется разное количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям.
Образование пыльцевых зерен и зародышевых мешков у цветковых растений
Изучая гаметогенез, нельзя не поражаться тому параллелизму, который наблюдается при созревании половых клеток у животных и растений, несмотря на то, что их расхождение (дивергенция) в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов как в растительном, так и животном мире.
Итак, изучение развития половых клеток у животных и у растений показало, что формирование гамет является сложным процессом. Прежде чем яйцеклетка и спермий объединятся в процессе оплодотворения, они претерпевают ряд превращений. Однако половые клетки так же, как и клетки любой другой ткани, происходят из соматических. Поэтому их нельзя рассматривать как нечто обособленное от тела организма. Вместе с тем половые клетки имеют и свои особенности. Основными характерными моментами, отличающими их от соматических клеток, являются следующие:
1. У разных животных и растений на разных стадиях дифференциации тканей зародыша происходит обособление половых клеток. Процесс закладки и дифференциации, половых клеток у животных называется зачатковым путем.
2. В процессе развития половых клеток особое значение имеет мейоз с характерными для него стадиями деления ядра, а именно профазой I, во время которой конъюгируют гомологичные хромосомы, метафазой I и анафазой I, когда осуществляется редукция числа хромосом и расхождение гомологичных хромосом к различным полюсам.
3. Главным свойством половых клеток является способность их при оплодотворении сливаться в одну с образованием зиготы, которая претерпевает затем дробление и развитие. Соматические клетки этой способностью, как правило, не обладают.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Развитие управляющих функций мозга ребёнка: полезные советы и упражнения для педагогов
Сертификат и скидка на обучение каждому участнику
. Микроспорогенез и формирование мужского гаметофита у покрытосеменных .
Микроспорогенез или образование мужского гаметофита. Взрослая тычинка состоит из пыльника, связника и тычиночной нити, которая одета эпидермой. Тычинка образуется из бугорка конуса нарастания цветоносного побега, который сначала однороден, а затем дифференцируется. Постепенно в двух участках каждого пыльцевого мешка начинается заложение гнезд пыльника; выделяется археспориальная клетка, а она делится, образуя париетальную и спорогенную клетки.
Париетальные клетки дают субэпидермальный фиброзный слой и выстилающий – тапетум, обеспечивающий спорогенную ткань питательными веществами. Цитоплазма густая, вязкая, клетки крупные.
Спорогенные клетки, делясь мейозом, дают клетки микроспор, в дальнейшем дифференцирующихся в пыльцу.
Ядро микроспоры делится на вегетативное и генеративное ядра.
Вегетативное - крупное, содержащее жирное масло и крахмал.
Генеративное - мельче, с хроматином.
Генеративное может делиться еще в пыльнике, образуя два спермия - (сложноцветные, злаки, маревые, гвоздичные), или же в пыльцевой трубке (орхидные, норичниковые).
Следовательно, зрелые пыльцевые зерна могут быть двух- и трехъядерные, покрыты спородермой, состоящей из экзины и интины Экзина - утолщенная слоистая оболочка, с выростами в виде шипиков, бугорков и т.д. Состоит из вещества - полленина, клетчатки, пропитанной кутиноподобными веществами, маслом с каротиноидами.
Интина состоит из пектиновых веществ.
Экзина имеет поры. Форма и характер поверхности экзины - диагностический признак.
Вскрывание пыльников обусловлено, в основном, сокращением клеток эпидермы при подсыхании.


Мегаспорогенез и формирование женского гаметофита у покрытосеменных.
Мегаспорогенез или формирование женского гаметофита. Семяпочки развиваются на внутренних стенках завязи и являются мегаспорангиями, заключенными в интегументы. Закладываются в виде бугорков на плаценте (место прикрепления семяпочки). Главная масса семяпочки - эмбриональные клетки, образующиеся из бугорков, формирующие нуцеллус (питательная и защитная ткань зародышевого мешка). Нуцеллус покрыт интегументами , оставляющими узкий канал в виде пыльцевхода (микропиле). Часть семяпочки, противоположная пыльцевходу – халаза.
Количество семяпочек в завязи неодинаково (слива, вишня - 1-2; злаки - 1; мак - много), соответственно этому неодинаково и число семян.
Форма зародышевого мешка разнообразна: овальный ,прямой, изогнутый и т.п. В цитоплазме зародышевого мешка имеются лейкопласты, хондриосомы, крахмальные зерна, капли масла и т.д.
Семязачаток – это небольшое образование, которое располагается в завязи пестика. Число семязачатков в завязи пестика может быть от одного (пшеница, ячмень, слива, подсолнечник) до нескольких тысяч (мак) и миллионов (орхидные). В семязачатке происходят процессы формирования мегаспор, женского гаметофита и процесс оплодотворения. После оплодотворения он развивается в семя.
Сформированный семязачаток представляет собой многоклеточное образование. К плаценте он прикрепляется семяножкой – фуникулюсом.
Снаружи семязачаток покрыт одним-двумя покровами – интегуметами. На верхушке интегументы не смыкаются, образуя таким образом канал – микропиле (пыльцевход) .
Под интегументами расположен многоклеточный нуцеллус , или ядро семязачатка (гомологичен мегаспорангию).
Конец семязачатка, противоположный микропиле, называют халазой . На уровне халазы происходит слияние нуцеллуса и интегументов (рис. 13).
Рис. 13. Строение сформированного семязачатка:
1 , 2 – внутренний и наружный интегументы; 3 – яйцеклетка; 4 – зародышевый мешок; 5 – нуцеллус; 6 – халаза; 7 – антиподы; 8 – вторичное ядро; 9 – синергиды; 10 – семяножка;
11 – плацента; 12 – проводящий пучок; 13 – пыльцевход (микропиле)
Особенности строения зрелого семени во многом определяются типом образовавшего его семязачатка. В зависимости от ориентации, взаимного расположения частей и степени изогнутости нуцеллуса различают пять основных типов семязачатков (рис. 14):

Рис. 14. Основные типы семязачатков:
1 – ортотропный; 2 – анатропный; 3 – гемитропный; 4 – кампилотропный;
5 – амфитропный
1) ортотропный (прямой) – микропиле и семяножка находятся на противоположных концах оси семязачатка (гречишные, ореховые);
2) анатропный (обратный, обращенный) – нуцеллус повернут по отношению к прямой оси на 180°, поэтому микропиле и семяножка расположены рядом (самый распространенный тип семязачатка);
3) гемитропный (полуповернутый) – семязачаток повернут на 90°, в результате микропиле и нуцеллус расположены по отношению к семяножке под углом 90° (некоторые первоцветные, норичниковые);
4) кампилотропный (односторонне изогнутый) – нуцеллус изогнут только микропилярным концом, фуникулюс и микропиле расположены рядом (бобовые, мальвовые);
5) амфитропный (двусторонне изогнутый) – нуцеллус изогнут с двух сторон, в виде подковы, микропиле и фуникулюс расположены рядом (тутовые, ладанниковые).
Мегаспорогенез – это процесс формирования мегаспор в нуцеллусе семязачатка.В нуцеллусе вблизи микропиле обособляется, увеличивается одна (реже несколько) клета археспория. Эта клетка становится материнской клеткой мегаспор, она имеет диплоидный набор хромосом (2n). Материнская клетка однократно делится мейозом , в результате возникает тетрада гаплоидных мегаспор (четыре по 1n). Мегаспоры располагаются линейно от микропиле к халазе (рис. 15).

Рис. 15. Развитие зародышевого мешка (схема):
1 – материнская клетка; 2 – тетрада мегаспор; 3 – одна развитая мегаспора и три отмирающие; 4 – двухъядерный мешок; 5 – четырехъядерный мешок; 6 – восьмиядерный мешок; 7 – развитый зародышевый мешок; с – синергиды; я – яйцеклетка;
п – полярные ядра; а – антиподы; м – три неразвитые мегаспоры; мз – мейоз;
Мегагаметогенез – это процесс формирования женского гаметофита, которым является восьмиядерный зародышевый мешок. Одна из четырех мегаспор, которая находится ближе к халазе, делится быстрее. Поэтому остальные три мегаспоры подавляются в развитии, т. е. дегенерируют.
Единственная мегаспора увеличивается, разрастается, ее ядро трехкратно делится митозом . В результате в сильно растянутой клетке мегаспоры формируется восемь ядер, которые располагаются по четыре на халазальном и микропилярном полюсах. Далее происходит дифференцировка ядер: от полюсов к центру отходит по одному ядру. Эти ядра называют полярными . Оставшиеся ядра обособляются в клетки. На микропилярном полюсе центральная крупная клетка преобразуется в яйцеклетку , две рядом расположенные боковые клетки – в синергиды . Яйцеклетка с синергидами образует яйцевой аппарат . На противоположном халазальном полюсе три клетки образуют группу антипод . В центре два полярных ядра сливаются, в результате образуется вторичное (центральное) ядро зародышевого мешка (см. рис. 13).
Таким образом, единственная мегаспора , прорастая внутри семязачатка, образует женский гаметофит – зародышевый мешок. Он включает:
а) яйцевой аппарат состоит из яйцеклетки и двух синергид; они расположены на микропилярном полюсе семязачатка;
б) три антиподы – расположены на халазальном полюсе семязачатка; яйцеклетка, синергиды и антиподы являются гаплоидными (1n);
в) вторичное (центральное) ядро зародышевого мешка – образовано при слиянии двух полярных ядер; оно диплоидное (2n).

Оплодотворение. После опыления начинается прорастание пыльцы. Липкая и неровная поверхность рыльца способствует удержанию большого числа пыльцевых зерен. Кроме того, если партнеры совместимы, рыльце выделяет специальный энзим, стимулирующий прорастание пыльцы. Прорастает несколько пыльцевых зерен. От индивидуальной скорости роста зависит успех той или иной пыльцы. В пыльцевую трубку переходят два спермия и вегетативное ядро, которое располагается у растущего конца трубки. Трубка входит в семяпочку через микропиле, конец ее лопается и содержимое изливается в полость зародышевого мешка. Один спермий сливается с гаплоидной яйцеклеткой, другой с диплоидной вторичной клеткой.
Обычно в зародышевый мешок проникает одна пыльцевая трубка, если семяпочек много, то несколько выполняют свое назначение.
Из диплоидной зиготы развивается зародыш семени, из триплоидной - эндосперм. Описанный универсальный для покрытосеменных процесс - двойное оплодотворение , открыт Сергеем Гавриловичем Навашиным в 1898 году.
От голосеменных отличия:
1) гибридный, триплоидный эндосперм;
2) два (а не один) спермий.
3) время от опыления до оплодотворения - 1-2 дня.
Смена ядерных фаз и чередование поколений . Женский гаметофит - зародышевый мешок.
Мужской гаметофит - пыльца.
Обыкновенный тип двойного оплодотворения - амфимиксис . Но у некоторых растений зародыш развивается из неоплодотворенных элементов. Отклонения от нормального хода оплодотворения - апомиксис . Различают:
1. Партеногенез –развитие зародыша из неоплодотворенной яйцеклетки;
2. Апогамия - развитие зародыша из антиподы или синергиды;
3. Апоспория - зародыш развивается из клетки нуцеллуса или интегумента.
После оплодотворения начинается деление оплодотворенных элементов внутри зародышевого мешка. Триплоидная зигота образует эндосперм, раньше зародыша. Зародыш развивается из зиготы. Может быть и перисперм - из нуцеллуса (вне зародышевого мешка). Интегументы становятся кожурой семени.
Зародыш, эндосперм, перисперм и кожура образуют семя .
Различают 4 типа семян.
1. Зародыш занимает всю полость зародышевого мешка, вытесняя весь эндосперм и перисперм. Питательные вещества - в семядолях (бобовые, тыквенные, розоцветные, сложноцветные).
2. Зародыш занимает меньшую часть семени, большая принадлежит эндосперму. Перисперма не образуется (злаки, пасленовые, зонтичные).
3. Есть только перисперм. Эндосперм полностью потреблен зародышем в процессе формирования (гвоздичные, маревые).
4.Сохраняется эндо- и перисперм - редкий тип семян (черный перец, лотос).
При образовании зародыша зигота делится на две клетки, образуя
1) подвесок или суспензор,
2) собственно зародыш.
Подвесок погружает зародыш в эндосперм, нередко выполняет функции присоски.
Зародыш состоит из зародышевого корешка, стебелька, к верхней части которого прикрепляются семядоли - первые листья растения. Стебелек ниже семядолей - гипокотиль, выше - эпикотиль; верхушка стебля - почечка.
Задание 27 № 11500 Какой хромосомный набор характерен для клеток зародыша и эндосперма семени, листьев цветкового растения. Объясните результат в каждом случае.
Пояснение. 1) в клетках зародыша семени диплоидный набор хромосом — 2n, так как зародыш развивается из зиготы — оплодотворённой яйцеклетки;
2) в клетках эндосперма семени триплоидный набор хромосом — 3n, так как образуется при слиянии двух ядер центральной клетки семязачатка (2n) и одного спермия (n);
3) клетки листьев цветкового растения имеют диплоидный набор хромосом — 2n, так как взрослое растение развивается из зародыша.
Задание 27 № 12073 Какой хромосомный набор характерен для ядер клеток эпидермиса листа и восьмиядерного зародышевого мешка семязачатка цветкового растения? Объясните, из каких исходных клеток и в результате какого деления образуются эти клетки. Пояснение. 1. Эпидермис листа имеет диплоидный набор хромосом. Взрослое растение является спорофитом.
2. Все клетки зародышевого мешка гаплоидны, но в центре находится диплоидное ядро(образуется в результате слияния двух ядер) — это уже не восьмиядерный, а семиклеточный зародышевый мешок. Это гаметофит.
3. Спорофит образуется из клеток зародыша семени путем митотического деления. Гаметофит образуется путем митотического деления из гаплоидной споры.
Задание 27 № 18711 Какой хромосомный набор характерен для вегетативной, генеративной клеток и спермиев пыльцевого зерна цветкового растения? Объясните, из каких исходных клеток и в результате какого деления образуются эти клетки.
Пояснение. 1) набор хромосом вегетативной и генеративной клеток — n;
2) вегетативная и генеративная клетки пыльцы образуются путём митоза при прорастании гаплоидной споры;
3) хромосомный набор спермиев — n;
4) спермии образуются из генеративной клетки путём митоза.
Задание 27 № 20388 В соматических клетках овса 42 хромосомы. Определите хромосомный набор и количество молекул ДНК перед началом мейоза I и в метафазе мейоза II. Ответ поясните.
Пояснение. 42 = 2 n — соматическая клетка, 21 = n — гамета, n — количество хромосом, с — количество молекул ДНК.
Перед началом мейоза I в интерфазе происходит репликация (удвоение) ДНК, клетка имеет диплоидный набор хромосом, но с удвоенными ДНК — 2 n 4 c , значит, клетки семязачатка в этот период имеют 42 хромосомы и 84 молекулы ДНК.
В конце мейоза I произошло редукционное деление (в каждую клетку отошло по одной хромосоме из пары гомологичных хромосом), после этого каждая клетка имеет имеет хромосомный набор 1 n 2 c . В метафазе мейоза II хромосомы выстраиваются по экватору клетки, хромосомный набор не изменяется, значит, в клетках в эту фазу 21 хромосома и 42 молекулы ДНК.
1) способы деления клеток при образовании микроспор из спорогенной ткани;
2) способы деления при образовании вегетативной и генеративной клеток покрытосеменного растения;
3) число хромосом и молекул ДНК в микроспоре, вегетативной и генеративной клетках покрытосеменного растения (выразить формулой). Пояснение. Элементы ответа.
1) Микроспоры образуются в результате мейоза.
2) Вегетативная и генеративная клетки образуются в результате митоза.
3) Во всех указанных клетках гаплоидный набор хромосом и ДНК – nc
Задание 27 № 21340 Какой хромосомный набор характерен для клеток восьмиядерного зародышевого мешка и зародышевой почечки семени пшеницы. Объясните, из каких исходных клеток и в результате какого деления они образуются.
Задание 27 № 21705 Определите число (n) хромосом и количество ДНК (с) у спор, заростка, половых клеток и спорофита папоротника. В результате какого деления образуются эти клетки и стадии развития?
Задание 27 № 22016 В соматической клетке кукурузы 20 хромосом. Определить набор хромосом в клетке верхушки растения и в ядре пыльцевого зерна.
Задание 27 № 22023 Какой хромосомный набор характерен для мегаспоры и клеток эндосперма сосны? Объясните, из каких исходных клеток и в результате, какого деления образуются эти клетки.
Хромосомный набор соматических клеток пшеницы равен 28. Определите хромосомный набор и число молекул ДНК в клетках кончика корня в метафазе и конце телофазы митоза. Объясните все полученные результаты.
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Навашиным в 1910 г. на лилейных растениях.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит — это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Схема мегаспорогенеза (1-5) и мегагаметогенеза (6-12) у растений
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры. В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других — в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения. Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно. В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других — из двух (биспорический тип) и четырех спор (тетраспорический тип).
Типы развития зародышевых мешков
Как мы указывали, при моноспорическом типе развивается лишь одна мегаспора из четырех, а остальные три разрушаются подобно тому, что имеет место с редукционными тельцами у животных. При других типах развития зародышевого мешка сохраняется разное количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям.
Образование пыльцевых зерен и зародышевых мешков у цветковых растений
Изучая гаметогенез, нельзя не поражаться тому параллелизму, который наблюдается при созревании половых клеток у животных и растений, несмотря на то, что их расхождение (дивергенция) в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов как в растительном, так и животном мире.
Итак, изучение развития половых клеток у животных и у растений показало, что формирование гамет является сложным процессом. Прежде чем яйцеклетка и спермий объединятся в процессе оплодотворения, они претерпевают ряд превращений. Однако половые клетки так же, как и клетки любой другой ткани, происходят из соматических. Поэтому их нельзя рассматривать как нечто обособленное от тела организма. Вместе с тем половые клетки имеют и свои особенности. Основными характерными моментами, отличающими их от соматических клеток, являются следующие:
1. У разных животных и растений на разных стадиях дифференциации тканей зародыша происходит обособление половых клеток. Процесс закладки и дифференциации, половых клеток у животных называется зачатковым путем.
2. В процессе развития половых клеток особое значение имеет мейоз с характерными для него стадиями деления ядра, а именно профазой I, во время которой конъюгируют гомологичные хромосомы, метафазой I и анафазой I, когда осуществляется редукция числа хромосом и расхождение гомологичных хромосом к различным полюсам.
3. Главным свойством половых клеток является способность их при оплодотворении сливаться в одну с образованием зиготы, которая претерпевает затем дробление и развитие. Соматические клетки этой способностью, как правило, не обладают.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
§ 32. Чередование способов размножения и поколений в жизненном цикле растений
*Образование половых клеток и оплодотворение у покрытосеменных
В § 29—3 вы познакомились с бесполым размножением растений, а сейчас более подробно рассмотрим половое размножение растений на примере покрытосеменных.
Вам уже известно строение и функции генеративного органа покрытосеменных — цветка. Рассмотрим процессы, протекающие в пыльнике тычинки и семязачатке завязи пестика.

Пыльник содержит пыльцевые гнезда (спорангии), в которых происходит образование спор. Из каждой материнской клетки путем деления образуется по четыре одноклеточные гаплоидные споры. Затем каждая спора делится на две клетки: крупную вегетативную и малую генеративную. Они покрываются двойной плотной оболочкой с порами, и образуется пыльцевое зерно (мужское половое поколение). После деления генеративной клетки формируются два спермия (мужские половые клетки без жгутиков). Все клетки в пыльцевом зерне гаплоидны.

Частью цветка, предопределяющей женский пол, является пестик . Он состоит из рыльца, столбика и завязи (см. рисунок в § 0—4). В завязи находится семязачаток (семяпочка), в котором снаружи имеются покровы. На верхушке семязачатка покровы не срастаются, и образуется пыльцевход. Одна из материнских клеток семязачатка вблизи пыльцевхода укрупняется, делится и образует четыре гаплоидные споры. Три из них погибают. Из четвертой споры после нескольких делений ядра и цитоплазмы образуется семь клеток. По три клетки с гаплоидным набором хромосом находится у каждого полюса, а между ними располагается крупная центральная диплоидная клетка. Одна из трех клеток у полюса возле пыльцевхода становится яйцеклеткой. Две соседние клетки называются клетками-синергидами, а три клетки на противоположном полюсе — клетками-антиподами. Образовавшаяся структура из семи клеток с яйцеклеткой представляет собой зародышевый мешок (женское половое поколение).
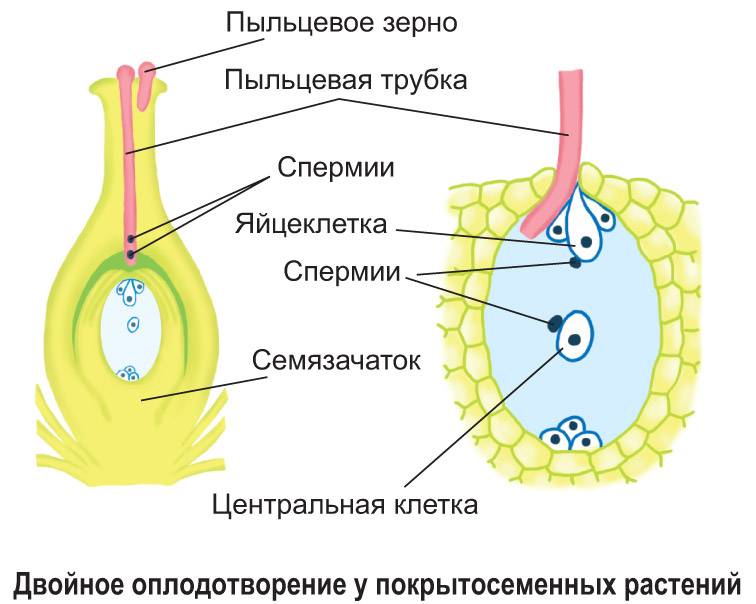
Перенос пыльцевого зерна из пыльника тычинки на рыльце пестика у покрытосеменных растений называется опылением. Оно может осуществляться с помощью насекомых, ветра, воды, птиц или самоопылением. После попадания на рыльце пестика пыльцевое зерно прорастает: вегетативная клетка вытягивается в длинную пыльцевую трубку, которая растворяет ткань столбика, проникает в полость завязи и через пыльцевход достигает зародышевого мешка. Спермии продвигаются по пыльцевой трубке, попадают в зародышевый мешок. Один из них оплодотворяет яйцеклетку зародышевого мешка, а второй сливается с центральной клеткой.
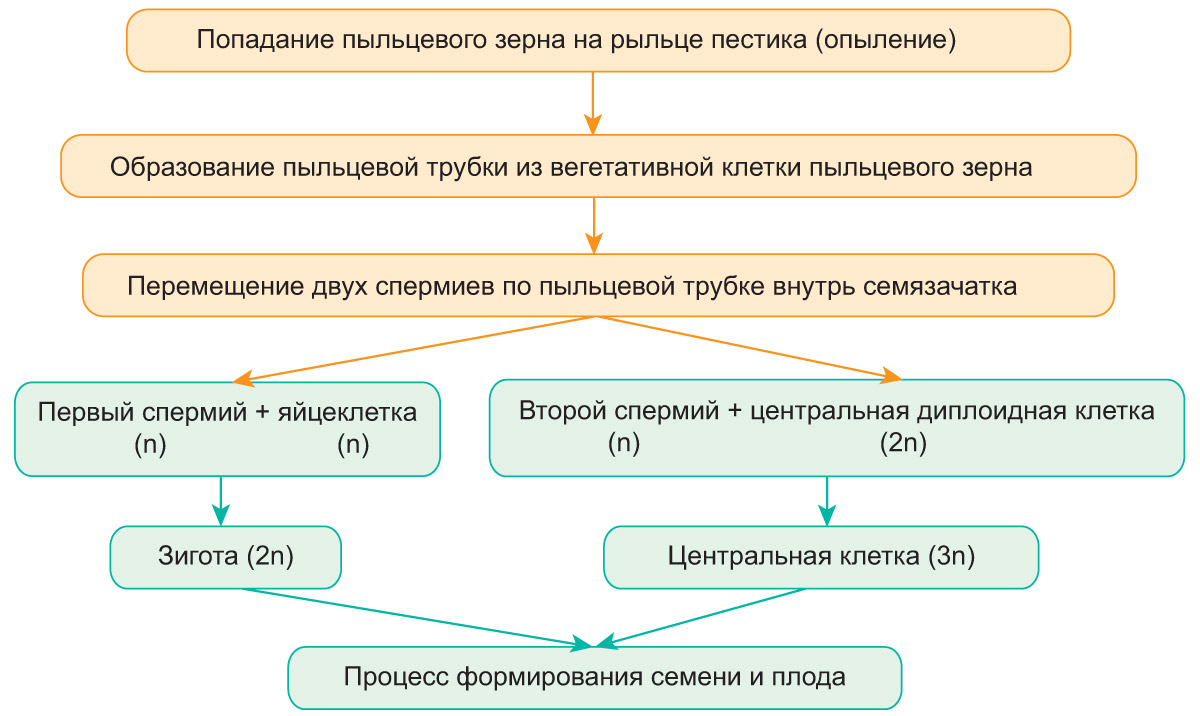
После образования зиготы в зародышевом мешке погибают синергиды. Центральная триплоидная клетка многократно делится, в результате чего образуется эндосперм, который содержит запас питательных веществ. Из зиготы формируется зародыш семени. После его образования антиподы погибают. Формирование семени и плода показано на схеме.
У некоторых растений в образовании околоплодника могут принимать участие чашечка и цветоложе.
Таким образом, семя образуется из семязачатка, а на месте завязи развивается плод.
Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных, где эндосперм развивается до оплодотворения, у покрытосеменных эндосперм образуется лишь в случае оплодотворения. Это обеспечивает существенную экономию энергетических ресурсов. Клетки эндосперма содержат триплоидный набор хромосом, что приводит к увеличению размеров клеток и количества питательных веществ, повышающих устойчивость зародыша к неблагоприятным факторам.
Читайте также: