
Спорогон у мхов это
Добавил пользователь Валентин П. Обновлено: 19.09.2024
МХИ, листостебельные мхи (Musci), класс отдела моховидных (Bryophyta) царства растений. Охватывает примерно 14 000 видов мелких споровых растений, встречающихся по всему миру в разнообразных средах, включая пустыни и водоемы, хотя большинство приурочено к влажным наземным местообитаниям.
Строение.
Многие мхи образуют прямостоячие неветвистые побеги, собранные в плотные дерновины или подушки. Другие, сильно ветвясь, стелются по почве или другим субстратам, например коре деревьев или скалам. Высота побегов обычно не превышает 5 см. Листорасположение спиральное; корней нет, а их функцию выполняют ризоиды.
Настоящие стебли и листья у мхов отсутствуют, и соответствующие им структуры обозначают особыми терминами – каулидии и филлидии (филлоиды). У многих мхов клетки каулидия недифференцированны, у других наблюдается центральный тяж из толстостенных клеток. Они, несомненно, выполняют опорную функцию, но не эквивалентны проводящей системе – специализированным тканям, служащим для транспорта воды и питательных веществ у сосудистых растений. Большая часть воды и солей, необходимых для жизнедеятельности мхов, втягивается в их тело из внешней среды капиллярными силами через щели между филлидиями и каулидием.
Филлидии бывают разными по форме и размеру. Обычно они состоят всего из одного слоя клеток, но у некоторых видов по краям филлидия этих слоев несколько. Если присутствует средняя жилка толщиной в несколько клеток, она бывает одиночной, доходящей до вершины филлидия, или двойной и короткой. У некоторых видов на ней образуются пластинчатые или столбчатые выросты. Форма филлидия бывает округлой, овальной, ланцетной, продолговатой или линейной, а его край может быть цельным или зубчатым, плоским или свернутым. Эти признаки достаточно видоспецифичны и используются в таксономии.
Ризоиды.
Роль корней играют многоклеточные ветвящиеся нити – ризоиды. У молодых мхов они всасывают из почвы воду с растворенными в ней минеральными веществами, но со временем эту способность утрачивают и служат просто для закрепления растения в субстрате.
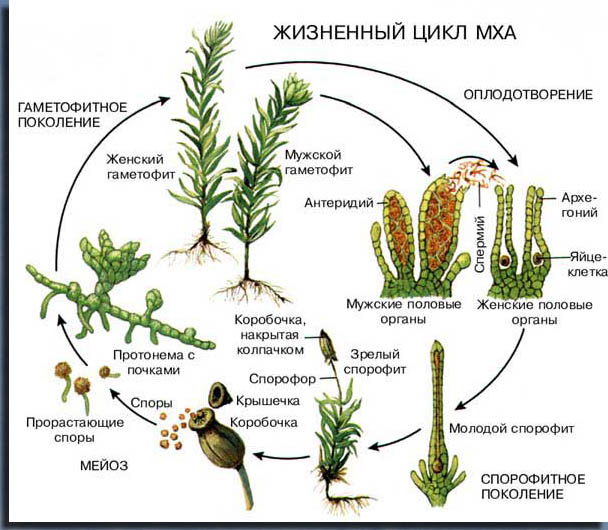
Жизненный цикл.
Зеленое фотосинтезирующее растение у мхов – это половое поколение, называемое гаметофитом. Гаметы, т.е. половые клетки, образуются на нем в особых половых органах (гаметангиях). Мужской гаметангий называется антеридием, женский – архегонием. Из оплодотворенной яйцеклетки (зиготы) развивается споровое поколение – спорофит. У мхов он практически лишен хлорофилла, остается прикрепленным к гаметофиту и получает от него питание. У спорофита каждая клетка содержит двойной (диплоидный) набор хромосом, а у гаметофита – одинарный (гаплоидный), как в гаметах. При слиянии спермия с яйцеклеткой из двух гаплоидных наборов образуется один диплоидный, необходимый для развития спорофита. У последнего при формировании спор происходит т.н. редукционное деление клеток (мейоз), каждая спора снова становится гаплоидной и может прорасти в такой же гаплоидный гаметофит.
Гаметофит.
Когда спора попадает во влажное место, она развивается сначала в ветвистую многоклеточную нить – протонему, или проросток. Оставшиеся на поверхности веточки протонемы становятся зелеными и фотосинтезирующими, а проникшие в грунт – бесцветными ризоидами. На зеленых частях проростка формируются боковые почки, из которых развиваются листостебельные побеги. Одна спора может дать целую колонию гаметофитов. У некоторых видов проростки долгоживущие, покрывающие иногда несколько квадратных дециметров грунта, у других – мелкие, исчезающие после появления облиственных побегов.
Спорофит.
Зигота начинает делиться еще в архегонии, который некоторое время растет вместе с формирующимся спорофитом. Когда он становится видимым невооруженным глазом, то состоит из трех частей: стопы, погруженной в брюшко архегония, тонкой ножки – спорофора и коробочки, где созревают споры. Растущий спорофит разрывает архегоний по кругу и выносит его верхнюю часть вверх в виде покрывающего коробочку колпачка (калиптры). Типичная зрелая коробочка – сложная структура, состоящая из урночки, крышечки и соединяющего их слоя специализированных толстостенных клеток – колечка. Набухая от воды, колечко отделяется от соседних частей коробочки и крышечка опадает, открывая устье урночки, которое бывает гладким или окруженным перистомом (околоустьем) из одного или двух концентрических рядов зубчиков. Эти зубцы либо плоские, либо несут от 4 до 64 поперечных гигроскопичных утолщений. Их число и форма относятся к важным таксономическим признакам мхов.
В зрелой коробочке находится множество свободных спор. Они выдуваются или вытряхиваются оттуда, разносятся ветром, водой или животными и, попав в благоприятные условия, прорастают.
Чередование поколений и эволюция.
Моховидные, или Мхи, или Настоящие мхи, или Бриофиты (лат. Bryophyta) — отдел высших растений, насчитывающий около 10 000 видов, объединённых примерно в 700 родов и 110—120 семейств (общее число всех мохообразных, включая Печёночные мхи и Антоцеротовые мхи, составляет около 20 000 видов). Как правило, это мелкие растения, длина которых лишь изредка превышает 50 мм; исключение составляют водные мхи, некоторые из которых имеют длину более полуметра, и эпифиты, которые могут быть ещё более длинными. Моховидные, как и другие Мохообразные, отличаются от других высших растений тем, что в их жизненном цикле гаплоидный гаметофит преобладает над диплоидным спорофитом.
Ранее в этот отдел, помимо класса листостебельных мхов, или собственно мхов (в ранге класса — Bryopsida), включали также Печёночные мхи (в ранге класса — Marchantiopsida, или Hepaticopsida) и Антоцеротовые мхи (в ранге класса — Anthocerotopsida), но сейчас эти таксоны повышены до уровня собственных отделов Marchantiophyta и Anthocerotophyta. Для обозначения совокупности этих трёх отделов используется неформальный собирательный термин мохообразные (Bryophytes).
Раздел ботаники, предметом изучения которого являются мохообразные, называется бриологией.
История.
Многие данные указывают на то, что мхи, как и другие споровые растения, произошли от псилофитов (риниофитов) — группы древних вымерших наземных растений. При этом спорофит мхов рассматривается как конечный результат редукции разветвлённого спорофита предковых форм.Есть и другая точка зрения, согласно которой мхи, плауновидные и риниофиты имеют общее происхождение от ещё более древней группы растений.Ранние палеонтологические находки датируются концом девона — началом карбона.
Биологическое описание
У мхов нет цветков, корней и проводящей системы. Размножаются мхи спорами, которые созревают в спорангиях на спорофите. В жизненном цикле, в отличие от сосудистых растений, преобладает гаплоидный (то есть с одинарным набором непарных хромосом) гаметофит (половое поколение). Гаметофит мхов — многолетнее зелёное растение, нередко с листоподобными боковыми выростами и корнеподобными выростами (ризоидами), в то время как спорофит (или бесполая стадия жизненного цикла) короткоживущий, быстро усыхает и состоит только из ножки и коробочки, в которой созревают споры.
Спорофит моховидных (носящий название спорогоний, или спорогон), имеет более простое строение, чем у других групп высших растений. Он не способен укореняться и располагается на гаметофите. Спорофит состоит, как правило, из трёх элементов:
- коробочка (или спорангия), в которой развиваются споры;
- ножка (или спорофора), на которой располагается коробочка;
- стопы, обеспечивающей физиологическую связь с гаметофитом.
Хлоропласты в клетках Plagiomnium affine из семейства Mniaceae
Со мхами не следует путать так называемый Олений мох (Ягель), поскольку олений мох — собирательное название для нескольких лишайников.
Распространение
Мох, покрывший камни
Мхи встречаются на всех континентах, в том числе в Антарктиде, нередко в экстремальных условиях обитания.
Обычно мхи образуют плотные скопления в затенённых местах, нередко в непосредственной близости от воды, хотя могут встречаться и на относительно открытых, сухих участках. Среди моховидных есть виды, живущие в пресноводных водоёмах. Морских обитателей нет, хотя некоторые виды поселяются на скалах в полосе прибоя.

Значение мхов
- Участвуют в создании особых биоценозов, особенно там, где почти сплошь покрывают почву (тундра).
- Моховой покров способен накапливать и удерживать радиоактивные вещества.
- Играют большую роль в регулировании водного баланса ландшафтов, так как способны впитывать и удерживать большое количество воды.
В деятельности человека:
- Могут ухудшать продуктивность сельскохозяйственных земель, способствуя их заболачиванию.
- Предохраняют почву от эрозии, обеспечивая равномерный перевод поверхностного стока вод в подземный.
- Некоторые сфагновые мхи применяются в медицине (в качестве перевязочных средств при необходимости).
- Сфагновые мхи — источник образования торфа.
Классификация
Листостебельные представители моховидных имеют ряд признаков, которые отличают их от остальных высших растений.

У них отсутствуют корни; прикрепление к субстрату (частично и всасывание воды) осуществляется одним или многочисленными ризоидами — выростами эпидермы. Поглощение воды происходит главным образом нижней частью стебля или телом всего растения (сфагновые мхи).Вегетативные органы сформированы ассимиляционной, проводящей, запасающей и покровной тканями. Однако в отличие от других высших растений у моховидных отсутствуют настоящие сосуды и механические ткани. На стебле зеленых мхов по спирали расположены бесчерешковые листья, имеющие цельную пластинку с жилкой или без нее.Это единственная группа растений, эволюция которой связана с регрессивным развитием спорофита и преобладанием в жизненном цикле гаметофита. Спорофит моховидных —спорогон — состоит из коробочки и ножки, нижняя расширенная часть которой — стопа -— внедряется в ткань гаметофита и доставляет спорофиту воду и питательные вещества. Спорофит, таким образом, лишен самостоятельности и полностью зависит от гаметофита.
Основная функция спорофита — образование спор, которые в благоприятных условиях прорастают. С момента прорастания споры начинается развитие гаметофита — полового поколения моховидных. Прежде всего развивается разветвленная нитчатая (у большинства мхов) или пластинчатая (у сфагнума) протонема, на которой закладываются почки. У одних мхов из почек протонемы формируются пластинчатые слоевища, у других — листостебельные побеги гаметофита (взрослого мха), на которых развиваются органы полового размножения — антеридии и архегонии.
Самую многочисленную группу моховидных составляет класс Настоящие мхи (около 10 тыс. видов), включающий зеленые и сфагновые мхи.
Зеленые мхи.
Они встречаются повсеместно: произрастают на почве, стволах деревьев, скалах и крышах домов, но наиболее широко распространены в сырых лесах, образуя сплошной зеленый ковер. Один из наиболее известных и самых распространенных мхов — кукушкин лен, образующий густые дерновинки из прямостоячих неветвистых стеблей, густо покрытых узкими линейно-ланцетными листьями.Архегонии и антеридии формируются на верхушках стеблей разных особей, которые обычно растут рядом. В антеридиях созревают двухжгутиковые сперматозоиды, в архегониях — по одной неподвижной яйцеклетке. Оплодотворение происходит во время дождя или при обильной росе, поскольку сперматозоиды могут подплыть к архегонию только в воде. Из зиготы здесь же, на верхушке женского гаметофита, развивается спорофит .Внутри коробочки спорогона размещается спорангий, в котором образуются гаплоидные споры. После созревания споры высыпаются из коробочки, разносятся ветром и в благоприятных условиях прорастают, образуя протонему в виде зеленой разветвленной нити. На ней закладываются почки, из которых со временем развиваются листостебельные растения мха.Зеленые мхи способны также к вегетативному размножению молодыми побегами, почками, листьями, выводковыми телами.

Сфагновые мхи.
К сфагновым (белым, или торфяным, мхам) относится примерно 350 видов, составляющих один род сфагнум. Они распространены по всему миру, но наиболее обильно представлены на верховых болотах, где господствуют над всеми остальными видами растений.Строение сфагновых мхов отличается рядом особенностей. У них отсутствуют ризоиды, поэтому вода с растворенными минеральными солями поступает непосредственно в клетки листа и стебля. Стебель гаметофита несет мутовки ветвей, густо покрытых листьями, которые на верхушке главной оси образуют розетку в виде головки почковидной формы. Некоторые боковые, более длинные веточки свешиваются вдоль стебля и плотно прилегают к нему, создавая своеобразную проводящую систему наподобие фитиля, по которому поднимается вода.
Листья сфагновых мхов не имеют средней жилки. Все они состоят из двух типов клеток: живых — ассимилирующих, длинных и узких, с хлоропластами, и мертвых (без протопласта), с утолщениями на стенках и порами. Подобные клетки без живого содержимого присутствуют и в стебле. Благодаря такому анатомическому строению стебля и листа сфагнум способен впитывать и удерживать большое количество воды, которая в 30— 40 раз превышает массу самого растения. Поэтому почва, где поселяются эти мхи, постепенно переувлажняется и заболачивается.
Антеридии и архегонии сфагнумов образуются либо на разных особях, либо на разных веточках одного и того же растения. После оплодотворения из зиготы формируется шаровидная коробочка на ложноножке, являющейся частью гаметофита. При вскрытии коробочки зрелые споры разлетаются на расстояние до 10 см и прорастают, образуя пластинчатую протонему с ризоидом. Из почки на протонеме вырастает гаметофит.
Особенностью сфагновых мхов является беспрерывное нарастание стебля верхушкой и отмирание нижней части. Однако полного сгнивания отмерших частей не происходит, так как в пере-увлажненной почве мало кислорода, а вследствие выделения сфагнумом специфических органических кислот создается кислая среда, в которой прекращается развитие почвенных микроорганизмов (бактерий, грибов и др.), разлагающих растительные остатки. В течение длительного времени происходит накопление значительных запасов органических веществ в виде торфа. Процесс образования торфа идет очень медленно: слой торфа в 1 см накапливается за 10 лет, толщиной в 1 м — за тысячу лет.

Общая характеристика. Моховидные — это древняя, самая примитивная группа высших растений, ныне живущих на Земле. Древнейшие ископаемые формы мхов известны с каменноугольного периода.
По оценкам разных специалистов, современные моховидные представлены 2.0—25 тыс. видов, распространённых на всех континентах. Особенно это относится к Северному полушарию, где на больших|больших площадях (на болотах, иногда в лесах|лесах) моховидные доминируют в растительном покрове|покрове. Эти растения предпочитают наиболее увлажнённые места|места, однако произрастают и на каменистых склонах гор, в пустынях, на сухих открытых скалах, в тундрах, но не встречаются в морях, на сильно засоленных почвах, в ледниках|ледниках и сыпучих песках.
Подавляющее большинство моховидных — низкорослые многолетние травянистые растения размером от 1 мм до нескольких сантиметров, реже до 60 см и более. У большинства из них тело расчленено на стебель и листья, но встречаются и слоевищные формы.
Листостебельные представители моховидных имеют ряд признаков, которые отличают их от остальных высших растений. У них отсутствуют корни; прикрепление к субстрату (частично и всасывание воды|воды) осуществляется одним или многочисленными ризоидами — выростами эпидермы. Поглощение воды|воды происходит главным образом нижней частью стебля или телом всего растения (сфагновые мхи).
Вегетативные органы|органы сформированы ассимиляционной, проводящей, запасающей и покровной тканями. Однако в отличие от других высших растений у моховидных отсутствуют настоящие сосуды и механические ткани. На стебле|стебле зелёных мхов по спирали расположены бесчерешковые листья, имеющие цельную пластинку с жилкой или без неё.
Это единственная группа растений, эволюция которой связана с регрессивным развитием спорофита и преобладанием в жизненном цикле гаметофита. Спорофит моховидных —спорогон — состоит из коробочки и ножки, нижняя расширенная часть которой — стопа -— внедряется в ткань гаметофита и доставляет спорофиту воду и питательные вещества. Спорофит, таким образом, лишён самостоятельности и полностью зависит от гаметофита.
Основная функция спорофита — образование спор, которые в благоприятных условиях прорастают. С момента прорастания споры начинается развитие гаметофита — полового поколения моховидных. Прежде всего развивается разветвлённая нитчатая (у большинства мхов) или пластинчатая (у сфагнума) протонема, на которой закладываются почки. У одних мхов из почек протонемы формируются пластинчатые слоевища, у других — листостебельные побеги|побеги гаметофита (взрослого мха), на которых развиваются органы|органы полового размножения — антеридии и архегонии.
Самую многочисленную группу моховидных составляет класс Настоящие мхи (около 10 тыс. видов), включающий зелёные и сфагновые мхи.
Зелёные мхи. Они встречаются повсеместно: произрастают на почве, стволах деревьев, скалах и крышах домов, но наиболее широко распространены в сырых лесах|лесах, образуя сплошной зелёный ковёр. Один из наиболее известных и самых распространённых мхов — кукушкин лён|лен, образующий густые дерновинки из прямостоячих неветвистых стеблей|стеблей, густо покрытых узкими линейно-ланцетными листьями.
Архегонии и антеридии формируются на верхушках стеблей|стеблей разных особей|особей, которые обычно растут рядом. В антеридиях созревают двухжгутиковые сперматозоиды, в архегониях — по одной неподвижной яйцеклетке. Оплодотворение происходит во время дождя или при обильной росе, поскольку сперматозоиды могут подплыть к архегонию только в воде. Из зиготы здесь же, на верхушке женского гаметофита, развивается спорофит
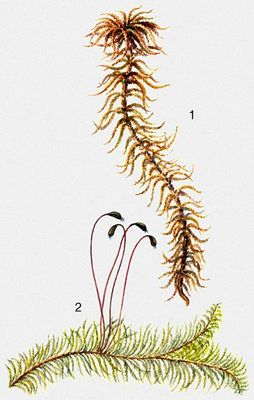
Мхи: а—кукушкин лён|лен; б—сфагнум; I —женское растение; 2 — мужское растение; 3 — женское растение со спорогоном; 4 — верхушка женского растения с архегониями (а); 5 — верхушка мужского растения с антеридиями (6); б — сперматозоид; 7— спорогон; 8 — спорогон на продольном разрезе; 9 — прорастание спор; 10 — протонема и основание стебелька; 11 — ассимилирующие и мёртвые водоносные клетки сфагнума; 12 — общий вид сфагнума.
Внутри коробочки спорогона размещается спорангий, в котором образуются гаплоидные споры. После созревания споры высыпаются из коробочки, разносятся ветром и в благоприятных условиях прорастают, образуя протонему в виде зелёной разветвлённой нити. На ней закладываются почки, из которых со временем развиваются листостебельные растения мха.
Зелёные мхи способны также к вегетативному размножению молодыми побегами, почками, листьями, выводковыми телами.
Сфагновые мхи. К сфагновым (белым, или торфяным, мхам) относится примерно 350 видов, составляющих один род сфагнум. Они распространены по всему миру, но наиболее обильно представлены на верховых|верховых болотах, где господствуют над всеми остальными видами растений.
Строение сфагновых мхов отличается рядом особенностей. У них отсутствуют ризоиды, поэтому вода с растворенными|растворёнными минеральными солями поступает непосредственно в клетки листа и стебля. Стебель гаметофита несёт мутовки ветвей, густо покрытых листьями, которые на верхушке главной оси|оси образуют розетку в виде головки почковидной формы. Некоторые боковые, более длинные веточки свешиваются вдоль стебля и плотно прилегают к нему, создавая своеобразную проводящую систему наподобие фитиля, по которому поднимается вода.
Листья сфагновых мхов не имеют средней жилки. Все они состоят из двух типов клеток: живых — ассимилирующих, длинных и узких, с хлоропластами, и мёртвых (без протопласта), с утолщениями на стенках и порами|порами. Подобные клетки без живого содержимого присутствуют и в стебле|стебле. Благодаря такому анатомическому строению стебля и листа сфагнум способен впитывать и удерживать большое количество воды|воды, которая в 30— 40 раз превышает массу самого|самого растения. Поэтому почва, где поселяются эти мхи, постепенно переувлажняется и заболачивается.
Антеридии и архегонии сфагнумов образуются либо на разных особях, либо на разных веточках одного и того же растения. После оплодотворения из зиготы формируется шаровидная коробочка на ложноножке, являющейся частью гаметофита. При вскрытии коробочки зрелые споры разлетаются на расстояние до 10 см и прорастают, образуя пластинчатую протонему с ризоидом. Из почки на протонеме вырастает гаметофит.
Особенностью сфагновых мхов является беспрерывное нарастание стебля верхушкой и отмирание нижней части. Однако полного сгнивания отмерших частей не происходит, так как в пере-увлажнённой почве мало|мало кислорода, а вследствие выделения сфагнумом специфических органических кислот создаётся кислая среда, в которой прекращается развитие почвенных микроорганизмов (бактерий, грибов и др.), разлагающих растительные остатки. В течение длительного времени происходит накопление значительных запасов органических веществ в виде торфа. Процесс образования торфа идёт очень медленно: слой торфа в 1 см накапливается за 10 лет, толщиной в 1 м — за тысячу лет.
В Беларуси болота занимают площадь около 2,5 млн. га с запасом торфа примерно 30 млрд. м3.
Торф находят широкое и разнообразное применение. Издавна его использовали как топливо и удобрение, особенно ценное для подзолистых почв. Мало|Мало разложенный торф в сельском хозяйстве служит для приготовления компостов, идёт на подстилку, помогает сохранению плодов и семян.
Торф является важным источником сырья для получения уксусной кислоты|кислоты, воска, парафина, нафталина, фенолов, спирта, карболовой кислоты|кислоты и т. д. Его используют в строительстве при изготовлении изоляционных плит, пластмасс и др.
Мхи, способные впитывать и удерживать большое количество воды|воды, играют существенную роль в регулировании водного баланса ландшафтов на значительной территории. Кроме того, они обеспечивают равномерный переход поверхностного стока вод в подземный, предохраняя почву от эрозии. Но в то же время при интенсивном развитии мхи могут заметно ухудшать продуктивность сельскохозяйственных земель и лугов, способствуя их заболачиванию.
Моховидные

Моховидные (мшистые) волокна|волокна мозжечка
Аксоныоливомозжечкового тракта распределены по всем отделаммозжечка и посылают коллатерали к глубиннымядрам мозжечка икоре мозжечка. Дистальные участки этих аксонов представляют собой особый типафферентных волокон мозжечка, так называемыелиановидные (лазящие) волокна|волокна мозжечка.
Все остальные афферентные пути оканчиваются в его коре моховидными(мшистыми) волокнами .
Мшистые волокна|волокна мозжечка возбуждают клетки-зерна|зёрна, а те через параллельные волокна|волокна оказывают возбуждающее действие на все остальные нейроны. Действие последних, однако, всегда является тормозным:клетки Гольджи тормозят клетки-зерна|зёрна по принципу обратной связи, а разряды клеток Пуркинье, возникающие под действием мшистых или лиановидных волокон, приводят к торможению нейронов ядер мозжечка.
Видео по теме : Моховидные
Все листостебельные мхи имеют более или менее развитое стеблевидное образование — каулидий, или стебель, покрытый листовидными выростами — листьями. У мхов установлены два основных типа роста стебля. Различают мхи ортотропные (от греч. orthos -прямой и tropos — поворот) с вертикальным ростом иплагиотропные (от греч. plagios — косой) с горизонтальным ростом стебля. Характер роста стебля хорошо согласуется с расположением на стебле гаметангиев, в соответствии с чем выделяют верхоплодные (акрокарпные) мхи и бокоплодные (плеврокарпные) мхи. К верхоплодным принадлежат мхи с прямостоячими стеблями, на верхушке которых образуются архегонии, а со временем и спорогон. Вторая группа объединяет мхи со стеблями, плоско прижатыми к субстрату, свисающими у эпифитных мхов, плавающими, как у фонтиналиса. У бокоплодных мхов архегонии закладываются на верхушке укороченных боковых побегов, и поэтому спорогоны сидят на боковых ветвях. Деление листостебельных мхов на верхоплодные и бокоплодные до некоторой степени является искусственным ввиду наличия переходных форм, но отражающим определенные тенденции в формативных процессах. В очертании на поперечном разрезе стебель чаще всего округлый, иногда овальный, угловатый или ребристый; последний образуется за счет сбегания жилки и краев основания листьев, которые, спускаясь по стеблю, срастаются с ним. Анатомическое строение стебля разнообразно. Стебель может быть построен из одинаковых клеток с толстыми или тонкими стенками или же из разных клеток. Во втором случае проявляется дифференциация на ткани: механическую, клетки которой по форме, утолщению стенок и их скульптуре сходны со склереидами сосудистых растений, и проводящие, обеспечивающие проведение воды и растворенных в ней минеральных веществ. Механическую ткань, которая составляет кору, или склеродерму, стебля мхов, образуют стереиды — механические клетки стебля. Это вытянутые в длину, узкие, проземхимные клетки, обычно окрашенные в желтоватый, бурый, красно-бурый, пурпурный или почти черный цвет. Как правило, у этих клеток толстые стенки, причем оболочка у них иногда настолько утолщена, что просвет почти полностью исчезает. Во внутренних утолщенных частях стенок имеются водопроводящие поры. На внутренней поверхности стенок стереидных клеток могут образовываться утолщения в виде папиллообразных выростов, образующих густую сеть в полости клетки. Они служат для сохранения воды (клеточная оболочка способна к набуханию). У ряда мхов один или несколько наружных слоев коры состоят из клеток, лишенных хлоропластов, с тонкими, прозрачными стенками и с более широким просветом. Эти слои клеток получили название гиалодермы (наружной коры). Гиалодерма развита у таких родов, как сфагнум, меезия (Meesia) и др. При подсыхании растения наружные стенки клеток гиалодермы впячиваются внутрь, а при поступлении воды стенки расправляются, клетки набирают воду и передают ее клеткам тканей, лежащих под гиалодермой. С внутренней стороны кора граничит с основной тканью, которая состоит из однородных паренхимных клеток, часто заполняющих всю внутреннюю часть стебля. Обычно кора постепенно переходит в основную ткань, реже кора резко отграничена от нее. Клетки основной ткани обильно заполнены плазматическим содержимым, хлоропластами, крахмалом, жирными маслами. Благодаря наличию хлоропластов основная ткань способна фотосинтезировать, но, кроме того, она служит местом сохранения воды, запасных веществ (крахмал, масла) и продуктов секреции (оксалат кальция). Основная ткань служит также 1 для проведения воды. У некоторых родов мхов в основной ткани обнаруживаются особые группы клеток, представляющие продолжение в стебле жилок листьев, по существу листовые следы. Различают два типа листовых следов: одни — слепо заканчивающиеся в основной ткани стебля и не доходящие до центрального пучка (обычно они состоят из группы одинаковых тонкостенных клеток), а другие — соединяющиеся с центральным пучком. Последние хорошо развиты в семействе политриховых (Polytrichaceae) в родах даусония (Dawsonia), сплахнум (Splachnum) и др. Листовые следы у политриховых имеют сложное строение, они дифференцированы на ткани, подобно стеблю этих мхов. Наличие или отсутствие центрального пучка в большинстве случаев является критерием для разграничения видов, а иногда и родов. Центральный пучок, проходящий в середине стебля, состоит из вытянутых в длину, большей частью тонкостенных клеток с узким просветом, обеспечивающих проведение воды или ее сохранение.
Наиболее сложно построен центральный пучок у рода политрихум (Polytrichum) и у других представителей семейства политриховых, а также у рода даусония. В стебле политриховых можно наблюдать ткани из клеток гидроидов и лептоидов, напоминающие ткани стебля примитивных сосудистых растений (особое сходство обнаруживается со стеблем риниевых). Гидроиды по характеру своей дифференциации, форме и отсутствию у них в зрелом состоянии цитоплазмы повторяют основные черты дифференциации и строения трахеид. По существу, единственное различие между теми и другими заключается в отсутствие у гидроидов лигнифицированных утолщений. Лептоиды, расположенные на периферии тяжа гидроидов, во многом сходны с ситовидными элементами папоротниковидных. Сходство это проявляется в удлиненной форме и закругленных концах, а также в наличии в концевых и боковых стенках многочисленных отверстий. В эпидерме стебля у всех мхов отсутствуют устьица. Кроме того, обычно из любой поверхностной клетки стебля могут возникать волосковидные образования -- ризоиды — однорядные многоклеточные нити с бурыми оболочками и с косыми поперечными стенками. Обычно они служат для прикрепления к субстрату и адсорбции воды. Развиваются ризоиды главным образом при основании прямостоячего стебля, как, например, у фунарии (Funaria), или по всей длине ползучего или лежачего стебля на брюшной стороне, обращенной к субстрату. Ризоиды иногда покрывают весь стебель густым, светлым или окрашенным в бурый, фиолетовый или красноватый цвет войлоком; у некоторых мхов они скручиваются в длинные тяжи. Ризоидный войлок обеспечивает поднятие из почвы воды в плотных дерновинках и более длительное ее сохранение в капиллярных пространствах между ризоидами и отдельными растениями. У водных мхов ризоиды выполняют главным образом функцию прикрепления к субстрату. На концах ризоидов у этих мхов образуются вильчатые разветвления, иногда сплетающиеся в подушечки, которыми растения прикрепляются к субстрату. При изменении условий питания и освещения ризоиды могут преобразовываться в зеленую вторичную протонему. На верхушке стебля, в пазухе листьев, в местах отхождения листьев от стебля очень рано возникают своеобразные нитевидные образования из нескольких клеток, которые называют булавовидными волосками. Их булавовидная конечная клетка способна выделять слизь для защиты точки роста стебля от высыхания. Кроме ризоидов и булавовидных волосков, на поверхности стебля у некоторых бокоплодных мхов развиваются пальчаторассеченные зеленые выросты стебля — парафиллии (от греч. para — возле и phyllon — лист). Иногда эти выросты образуются в таком количестве, что одевают стебель густым зеленым войлоком. Форма парафиллии чрезвычайно разнообразна. Они никогда не имеют жилок и расположены на стебле беспорядочно. Так же как и ризоиды, парафиллии способны не только проводить и удерживать воду, но и выполнять функцию фотосинтеза.
Ветвление у мхов не бывает дихотомическим. Оно обычно боковое, но не пазушное, поскольку каждая новая ветвь появляется ниже листа. У верхоплодных мхов стебель прямой или почти прямой и обычно слабоветвистый. У большинства видов ветви развиты только в верхней части стебля. Архегонии, а позднее и спорогоны возникают у них на верхушке стебля или основных ветвей; на развитие гаметангиев обычно тратится верхушечная клетка, и поэтому апикальный рост стебля и ветвей прекращается. У этих мхов, как правило, новые боковые ветви (инновации) образуются под гаметангиями и растут в направлении главного стебля, а на верхушке могут снова давать гаметангии. При симподиальном типе ветвления эти побеги возникают по одному; при вильчатом ветвлении или ложной дихотомии образуются парные побеги; у некоторых мхов возникает по нескольку побегов, т. е. имеет место пучковидное ветвление. При отмирании главного стебля такие инновации укореняются и становятся самостоятельными растениями. У бокоплодных мхов стебли стелющиеся (реже прямостоячие) и обильно моноподиально ветвящиеся. Архегонии и антеридии возникают у них не на верхушке главного стебля, а на верхушках укороченных боковых веточек вдоль главного стебля. Благодаря этому рост главного стебля при спороношении не приостанавливается и мох может расти дальше, достигая значительной длины. На каждой стороне стебля многочисленные ветви появляются в восходящей (акропетальной) последовательности, т. е. более молодые находятся ближе к верхушке. Образующиеся ветви обладают ограниченным верхушечным ростом. При правильном чередовании в развитии зачатков ветвей формируется перистое ветвление стебля; при повторном ветвлении — дважды-, триждыперистое ветвление, например у туидиума. Рост побегов и развитие ветвей как у верхоплодных, так и у бокоплодных мхов находится в зависимости от точки роста главного стебля. Так, если удалить верхушку стебля, то в новые побеги развиваются покоящиеся почки.
Листья у мхов сидячие, большей частью поперечно прикрепленные к стеблю, простые, цельные, по краю могут быть зубчатыми и только очень редко глубоко разделенными. На стебле листья расположены всегда по спирали, образуя двурядное или многорядное листорасположение. Закладываются листья в восходящей последовательности. Рост их начинается с деления двусторонней верхушечной клетки, от которой отделяются сегменты с двух сторон. После прекращения верхушечного роста листья достигают конечного размера посредством вставочного роста, происходящего в их основании. По расположению на стебле различают листья низовые, срединные (или собственно стеблевые) и покровные. Покровные листья окружают гаметангии я обычно отличаются от стеблевых размерами, формой, часто анатомическим строением, а иногда — окраской и консистенцией. Листья, окружающие архегонии, носят название пе-рихециальных, а листья, окружающие антеридии,— перигониалышх. Низовые листья — сильно редуцированные, чешуевидные. Они развиваются в нижней надземной или в подземной части стебля у таких мхов, снабженных ризомом, как климациум (Climacium). Стеблевые листья расположены выше низовых, преимущественно в средней и верхней частях стебля. Листовая пластинка чаще однослойная, реже целиком или частично дву-, многослойная. Клетки пластинки обычно богаты хлоропластами и выполняют функцию фотосинтеза. Жилка, если она развита, обычно проходит посередине листа и состоит из толстостенных, вытянутых клеток. Кроме механической роли, она обеспечивает проведение пластических веществ и воды. По форме листья очень разнообразны. Чрезвычайно различно строение краев листьев. Заварнутость и отвернутость краев листа создает микроскопические полости, где и удерживается капиллярная вода. Завернутый край листа, свойственный мхам, обитающим в условиях периодической засухи, выполняет еще и защитную функцию, предохраняя нежные части листа от иссушения. В листовой пластинке различают клетки двух основных типов: клетки паренхиматические — обычно округлые или угловатые до многоугольных (часто квадратные и шестиугольные), почти равной длины и ширины, и прозенхиматические — узкие, вытянутые в длину с заостренными, заходящими друг за друга концами. У мхов вся пластинка листа редко бывает построена из одинаковых клеток. Если верхняя часть листа состоит из паренхиматических-мелких клеток, более толстостенных, а иногда мамиллозных или папиллозных, то клетки листового основания почти всегда несколько крупнее и резко отличаются от верхних по форме. Большой полиморфизм клеток мхов обусловлен не только разнообразием их формы, но и характером их клеточных оболочек. Здесь их многообразие, кажется, не знает пределов. Кроме разрастания боковых стенок клеток пластинки листа, могут разрастаться и наружные стенки, образуя мамиллозность и папиллозность. Папиллы (утолщения клеточной оболочки) встречаются наиболее часто у бриевых мхов. Они бывают чрезвычайно разнообразной формы, а иногда к тому же в месте их образования клеточная оболочка разрастается и выпячивается. На этом образовании и сидят папиллы. Папиллы сильно увеличивают поверхность клеток, всасывающих воду, способствуют более быстрому собиранию воды и проведению ее внутрь клеток. Особенно часто развиваются папиллы на клетках у мхов сухих и сильно освещенных мест (тортула — Tortula, барбула — Barbula и др.), но широко представлены и у видов, обитающих в условиях избыточной влажности (у гелодиумашерстистого — Helodium lanatum, аулакомниума болотного — Aulacomnium pa-lustre, у многих лесных мхов из семейства туидиевых —Thuidiaceae, у типичных эпифитов тропических и субтропических лесов из семейства метеориевых — Meteoriaceae). У ряда мхов проявляется четкое обособление ассимиляционных и водоносных элементов листа. Это характерно для разных систематических групп мхов, далеко отстоящих друг от друга в родственном отношении. Наиболее ярко выраженный диморфизм клеток хорошо известен у сфагновых мхов, а из других листостебельных мхов — только у представителей семейства леукобриевых (Leucobryaceae) и калимперовых (Calymperaceae). Лист состоит из двух типов клеток: из зеленых, содержащих хлоропласты, и из мертвых, бесцветных, лишенных хлоропластов водоносных клеток. У сфагновых мхов хлорофиллоносные клетки развиты в большом количестве, окружают водоносные на протяжении всего листа. Водоносные клетки имеют спиральные и кольчатые утолщения и поры — отверстия в наружных оболочках. У леукобриевых в средней части многослойного листа расположено от одного до трех слоев хлорофиллоносных клеток. Почти у всех видов они со всех сторон окружены водоносными клетками с небольшим количеством пор преимущественно в основании листа. По экологии и географии две названные группы мхов сильно отличаются. Сфагновые мхи — типичные обитатели болот,
Домашние и дикие растения, их описание и выращивание; болезни и вредители. Интересные сведения из ботаники и других областей науки. Доклады для школьников. Природа Приморского края (растения и животные). История, культура, достопримечательности Владивостока и России в целом.
Вечерний Владивосток / улица Адмирала Фокина

- Главная страница
- Всё о бегониях
- Всё о бегониях - 2
- Список бегоний - 1
- Список бегоний - 2
- Список бегоний - 3
- Список бегоний - 4
- Список бегоний - 5
- Список бегоний - 6
- Список бегоний - 7
- Список бегоний - 8
- Список бегоний - 9
- Мои растения
- Доклады
- В помощь цветоводам, садоводам и огородникам
- Опознавалка бегоний и других растений
- Дизайн в саду и дома
- Комнатные растения
- Садовые растения
- Растения по алфавиту
- Растения по алфавиту (2)
- Приморский край
- Приморский край - 2
- Латынь в ботанике
- Биология
- Ботаники
- История и культура
- Интересное
- Вопрос-ответ
воскресенье, 10 мая 2020 г.
Спорангии и спорогенез у мхов

В простейшем случае, например у мхов , спорофит представлен небольшим диплоидным телом, на котором и развивается одиночный спорангий . Зигота, образующаяся в архегонии гаметофита , не теряет с ним связи и формирует спорофит , который фактически паразитирует на гаметофите .


Спорофит мхов обычно развивает более или менее длинную голую безлистную ножку, на верхушке которой и располагается спорангий .
Спорангий мхов представляет в большинстве случаев бокальчатую многослойную структуру, заполненную диплоидной спорогенной тканью, из которой и развиваются посредством мейоза гаплоидные споры .
Спорангий мхов имеет довольно сложное устройство. У одних мхов его верхушка имеет открывающуюся крышечку - оперкулум . Отверстие спорангия прикрыто, кроме опадающей крышечки, тонкой мембраной - эпифрагмой , а по краям несёт венчик зубчиков ( перистом ).
При подсыхании в сухую погоду эпифрагма и зубчики перистома отгибаются и позволяют спорам при покачивании спорангия свободно рассеиваться.
Вскрывшаяся коробочка у всех зелёных мхов имеет окраину - перистом ; он может быть простым или двойным. Типы перистома как по строению, так и по характеру образования весьма многочисленны. Перистом может иметь вид зубчиков, ресничек, сеточки; состоять из одной или нескольких клеток или только из стенок клеток кольца. Общим для перистома всех зелёных мхов является его гигроскопичность; во влажную погоду перистом набухает и занимает горизонтальное положение, стягивая к центру края коробочки. Зубчики перистома налегают на эпифрагму и закрывают коробочку целиком.
У видов, лишённых эпифрагмы , зубчики перистома длинные, и, смыкаясь своими вершинами, они также закрывают вход в урночку. Следовательно, перистом препятствует попаданию влаги внутрь коробочки и предотвращает слипание и загнивание спор .
Читайте также: