
Систематика характеристика особенностей строения и размножения подкласса зеленые мхи bryidae
Добавил пользователь Alex Обновлено: 15.09.2024
Подкласс бриевых наиболее обширный и полиморфный из всех листостебельных мхов. В нем насчитывают около 14 000 видов и более 700 родов, относящихся к 85 семействам. Это многолетние или однолетние мхи, различные по величине, от чрезвычайно мелких (высотой около 1 мм) до крупных (высотой до 50 см и более), обычно зеленые, реже красно-бурые: бурые, иногда черные.
Растут бриевые отдельными стебельками, группами, подушечками, дерновинками, ковриками или сплошными зарослями на различных субстратах — почве, коре стволов и ветвей, гнилой древесине, на содержащих известь и кислых горных породах, реже на разложившихся трупах мелких животных, на экскрементах животных, в местах различной степени увлажнения и освещения.
Распространены более широко, чем сфагновые мхи. Живут в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания бриевых мхов, где они доминируют или формируют сплошной покров, - тундра, болота и некоторые типы лесов. Каждому местообитанию свойственны свои виды. Бриевые мхи по сравнению со сфагновыми отличаются большим разнообразием строения. Органы полового размножения закладываются у одних видов на главной оси, у других - на боковых. У некоторых видов ветвление не выражено.
Политрих обыкновенный, кукушкин лен (Polytrichum commune) - это один из распространенных представителей бриевых мхов. Он растет в лесу, на полянах, окраинах болот.
Стебель гаметофита прямостоячий, неветвистый, высотой 15 см и более, густо покрыт листьями. Подземная часть его простирается в почве почти горизонтально, на ней образуются ризоиды. В центре стебля находится концентрический проводящий пучок, состоящий из вытянутых клеток, сходных с трахеидами и ситовидными трубками. Он окружен паренхимой, также выполняющей проводящую функцию. С наружной стороны паренхима граничит со склеродермой (корой). Внешний слой ее, состоящий из бесцветных клеток, называют гиалодермой.
Листья расположены по спирали. Они состоят из линейной пластинки с заостренной зубчатой верхушкой и пленчатого влагалища. На верхней стороне листа расположены ассимиляционные пластинки. Жилка с механическими и проводящими элементами расширена.
Гаметофит двудомный. Архегонии бутылковидной формы расположены на верхушке женского гаметофита, антеридии мешковидной формы - на верхушке мужского. Между архегониями и антеридиями имеются стерильные нити - парафизы. После оплодотворения из зиготы образуется спорогон, состоящий из длинной ножки и коробочки. Коробочка прямостоячая или более или менее косо расположенная, призматическая, четырех-пятиграниая, покрыта ржаво-войлочным колпачком, образующимся из стенок архегония. Коробочка состоит из урночки и крышечки. Нижняя часть урночки сужена в шейку. На границе урночки и шейки в эпидерме имеются устьица. В центре урночки расположена колонка, которая у крышечки расширяется и формирует эпифрагму - тонкостенную перегородку, закрывающую урночку. Вокруг колонки расположен спорангий в виде цилиндрического мешка, прикрепленного к стенке и колонке особыми нитевидными образованиями. Урночка имеет специальное приспособление для рассеивания спор - перистом, представляющий собой ряд зубчиков с тупыми верхушками, расположенных по краю урночки. Между зубцами, способными к гигроскопическим движениям, и эпифрагмой имеются отверстия, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На ней формируются почки, из которых со временем развиваются взрослые гаметофиты. Спорогон поднимается над перихециальными листьями или погружен в них. Коробочка обычно с перистомом, колечком и крышечкой, реже клейстокарпная. Споровый мешок пронизан колонкой (отсутствует у видов рода архидиум) и окружен, воздушной полостью. Споры большей частью округлые, гладкие или со сложной скульптурой, одноклеточные, редко многоклеточные, диаметром от 5 до 200 мкм у архидиума и некоторых родов гигаспермовых (Gigaspermaceae), чаще же 10-20 мкм.
Прорастание спор, образование протонемы и молодых растений у бриевых мхов происходит по особому типу. При прорастании спора набухает, затем эндоспорий, прорывая экзоспорий, вытягивается в нить. Эта нить ветвится, и ее ветви обладают ограниченным верхушечным ростом. Ризоиды тонкие, с бурыми стенками, косыми поперечными перегородками и с относительно немногочисленными хлоропластами или лейкопластами.
4. Класс Печеночники. Общая характеристика. Особенности морфологии, анатомии и размножения на примере маршанции многообразной.
Общее число видов около 10 тыс. Распространелы повсеместно. Примитивность строения тела печеночников свидетельствует об их древности. Маршанция обыкновенная (Marchantia polymorpha) - типичный представитель класса. Гаметофит в виде пластинчатого таллома, длиной 10 - 12 см, ветвление верхушечное. С обеих сторон он покрыт эпидермой. Верхняя эпидерма имеет вентиляционные отверстия - устьица. Они окружены специальными клетками, расположенными в четыре ряда. Под устьицами имеются воздушные камеры. Нижняя эпидерма дает выросты - одноклеточные ризоиды и красноватые или зеленоватые чешуйки, которые иногда принимают за редуцированные листья. Под верхней эпидермой расположена ассимиляционная ткань, состоящая из вертикальных столбиков паренхимных клеток с хлоропластами. Ниже находится слой тонкостенных бесхлорофилльных паренхимных клеток. Следовательно, таллом маршанции имеет дорсивентральное строение.
На верхней стороне таллома образуются особые веточки - подставки, а на них - органы полового размножения. Маршанция - двудомное растение. На одних экземплярах подставки имеют форму сидящей на ножке девятилучевои звездочки, между лучами которой на нижней стороне расположены архегонии. На других - подставки имеют форму сидящего на ножке восьмиугольного щитка, на верхней стороне которого расположены антеридии, погруженные в антеридиальные полости. В брюшке архегония образуется яйцеклетка. После ее слияния со сперматозоидом из зиготы образуется спорогон. Он представляет собой коробочку на короткой ножке, которая прикреплена к гаметофиту гаусторией. Внутри коробочки из спорогенных клеток в результате мейоза образуются гаплоидные споры, а также элатеры - мертвые удлиненные клетки со спирально утолщенной стенкой, служащие для разрыхления массы спор, а также для выбрасывания их из коробочки. В благоприятных условиях из споры развивается предросток, или протонема. Это небольшая нить. Из ее верхушечной клетки вырастает таллом маршанции.
Вегетативное размножение осуществляется выводковыми тельцами линзовидной формы, имеющими зеленый цвет. Они образуются на верхней стороне таллома в особых корзиночках в результате деления клеток, выстилающих их дно.
.Виды маршанции имеют широкое распространение. Наиболее часто их можно встретить во влажных местах: на берегах озер и рек, по оврагам и в травянистом покрове под пологом леса.
5. Отдел Плауновидные. Общая характеристика и классификация. Время наибольшего расцвета. Происхождение листьев плауновидных (микрофиллия). Цикл воспроизведения.
Плауновидные относятся к группе наиболее древних представителей земной флоры. В конце палеозойской эры, особенно в каменноугольном периоде, плауновидные достигли своего наибольшего развития и многообразия. В то время предки современных плауновидных, называемые сигилляриями, лепидодендронами или чешуедревами, покрывали большие просторы нашей планеты. В начале мезозойской эры, в связи с изменением климата, все крупные плауновидные начинают вымирать, их вытесняют более приспособленные к новым экологическим условиям древние голосеменные.
Мелкие травянистые виды плауновидных, выживающие под пологом более конкурентоспособных голосеменных, пережили их господство и успешно приспособились к дальнейшей жизни в новых экосистемах, где преобладающими растениями стали цветковые.
1400 видов современных плауновидных, имеющих совсем небольшое сходство со своими гигантскими предками, — это вечнозеленые многолетние травы, реже полукустарники. Очень редко они поднимают свои побеги, не имеющие камбия, выше чем на один метр. С помощью стелющихся побегов плауновидные хорошо и быстро вегетативно размножаются. При этом побеги, вырастающие на много метров в длину, создают впечатление ползучего или плывущего по почве растения. Не случайно этот большой отдел древних споровых растений получил название Плауновидные.
Побеги большинства растений этой группы покрыты спирально расположенными мелкими листьями – микрофиллами. Характерно для плауновидных также дихотомическое или вильчатое ветвление побегов. Подземные части этих растений обычно представлены корневищем с придаточными корнями и видоизмененными листьями. У некоторых видов подземные части побегов образуют своеобразный орган, несущий расположенные по спирали корни и называемый ризофором (от греч. rhiza – корень и phoros – несущий), или корненосцем. Надземные и подземные побеги плауновидных нарастают ограниченное время, поскольку со временем клетки верхушечных меристем теряют способность к делению.
Спороносные листочки (спорофиллы) плауновидных по форме, размерам и цвету могут быть похожи на вегетативные листья (трофофиллы) или в той или иной степени отличаться от них. Чередуясь со стерильными листьями, спорофиллы образуют на протяжении стебля спороносные зоны или собраны в расположенные на концах ветвей компактные образования – стробилы. У некоторых ископаемых форм плауновидных спорофиллы сидели на стебле вперемежку с вегетативными листьями, не образуя ни спороносных зон, ни стробилов.
Среди плауновидных есть равно- и разноспоровые растения, гаметофиты (заростки) которых, соответственно, либо обоеполые (т.е. на них образуются как мужские, так и женские половые клетки), либо однополые. Есть между ними и другие отличия. Заростки современных равноспоровых (представителей класса плауновых) созревают в течение 1–15 лет, достигая в длину от 2 до 20 мм. При этом они развиваются в подземных или полуподземных условиях, и питаются в значительной степени или исключительно за счет образования союза с почвенными грибами. Заростки разноспоровых плауновидных (к ним относятся все представители класса полушниковых) развиваются обычно в течение нескольких недель. Они вообще лишены хлорофилла и живут за счет питательных веществ, содержащихся в споре. Поэтому они очень мелкие и даже в зрелом виде лишь слегка выступают за пределы оболочки споры.
Половые органы, развивающиеся на заростках плауновидных, представлены антеридиями, в которых развиваются сперматозоиды с двумя или многими жгутиками, и архегониями, в которых развиваются яйцеклетки. Для того чтобы сперматозоид мог достичь яйцеклетки, необходимо наличие капельно-жидкой воды.
Надо заметить, что для того чтобы наблюдать эту фазу в цикле развития плаунов, как и других высших споровых растений, нужно проявить наблюдательность и терпение натуралиста-исследователя – разыскать крошечные заростки под травяно-кустарничковым ярусом леса очень непросто.
Оплодотворенная яйцеклетка у плауновидных сразу же, не впадая в состояние покоя, развивается в растение нового бесполого поколения – спорофит.
Все плауновидные никогда не образуют цветков — это споровые растения. Спорангии — органы спороношения — образуются в пазухах листьев или на их внутренних поверхностях. Споры могут созревать довольно долго, высыпаясь, они прорастают и дают начало гаметофитам только на влажных почвах.
Плауновидные, являясь обязательными компонентами многих экосистем, нуждаются в охране и защите. При заготовке спор, которые находили раньше широкое применение и используются сейчас в медицине и в некоторых отраслях промышленности, рекомендуется аккуратно отрезать только спороносные части растений, так называемые колоски, ни в коем случае не повреждать и не вырывать остальные части растения.
6. Начертить схему циклов воспроизведения плауна булавовидного и щитовника мужского. Укажите их сходство и различия.
У плаунов в жизненном цикле происходит чередование двух поколений, полового и бесполого.
То растение, которое мы видим в лесу– это спорофит (бесполое поколение). И размножается спорофит плауна булавовидного бесполым путем – спорами, мельчайшими пылинками без малейшего запаса питательных веществ. Размножение это очень ненадежно. Ведь из десятков тысяч спор попадет в благоприятные условия, прорастет и выживет, возможно, одна. И лишь огромное количество спор все же позволяет достичь результата.
Но попасть в благоприятные условия – это еще полдела! Некоторые споры обладают длительным периодом покоя и прорастают подземным способом. Такая спора может покрыться минеральными частичками почвы, может быть перемещена вглубь почвы водой, дождевыми червями, насекомыми. И там, в глубине, через несколько лет (от 3 до 8!) она прорастет.

Цикл развития плауна булавовидного: А — спорофит; Б — гаметофит; 1 — ползучий побег с придаточными корнями; 2 — восходящие побеги; 3 — ножка спороносных колосков; 4 — листочки: восходящего побега (а) и ножки спороносныхколосков (б); 5 — спороносные колоски; 6 — споролистики: вид с брюшной (в) и спинной (г) сторон; 7 — спорангии; 8 — споры; 9 — прорастающая спора; 10 — архегоний; 11 — антеридий; 12 — оплодотворение; 13 — оплодотворенная яйцеклетка;

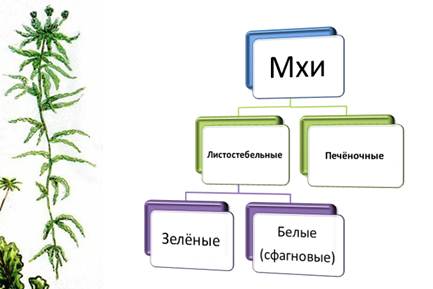
Листостебельные, или настоящие мхи, можно, в свою очередь, разделить на 3 подкласса:
- Бриевые (или зеленые) мхи (Bryidae);
- Сфагновые (или белые) мхи (Sphagnidae);
- Андреевые (или черные) мхи (Andreaeidae).
Класс антоцеротовые

Рис. 1. Антоцеротовые: а) антоцерос (Anthoceros laevis) — общий вид со зрелыми спорангиями; б) спорангий со спорами; в) архегоний; г) антеридий (внутри развивающиеся сперматозоиды); 1 — брюшко архегония (в центре яйцеклетка); 2 — шейка (внутри шейковые канальцевые клетки)
Устьица в эпидерме спорофита антоцеротовых состоят из двух замыкающих клеток и внешне похожи на устьица сосудистых растений. Спорофит антоцеровых, в отличие от спорофитов других моховидных, длительное время сохраняет способность к росту и фотосинтезу; показано, что он вообще может расти и питаться самостоятельно, без помощи гаметофита. Эти особенности антоцеротовых позволяют некоторым ученым видеть в них редуцированные сосудистые растения или даже низшее звено их эволюции (т.е. считать антоцеротовые возможными предками первых сосудистых растений — риниофитов).
Антоцеротовые очень сильно отличаются от других моховидных и вообще от других наземных растений. Печеночники и листостебельные мхи отличаются друг от друга значительно меньше. Возможно, происхождение антоцеротовых и других моховидных различно и их вообще следует считать различными отделами царства рас гения.
Во внутренних, заполненных слизью полостях гаметофита антоцеротовых обитают цианобактерии рода носток (Nostoc), которые фиксируют атмосферный азот и снабжают соединениями азота растение-хозяина. Однако медленнорастущие атоцеротовые- слабые конкуренты, поэтому большинство видов этого класса — обитатели нарушенных мест обитания (незадернованных краев пашен, канав, дорог, берегов рек).
Класс печеночники
Класс печеночники (Hepaticopsida), или печеночные мхи, объединяет около 10 000 видов.
Печеночники были названы так из-за того, что их слоевище своими очертаниями напоминает печень, поэтому в Средние века эти мхи считали эффективным средством для лечения ее заболеваний.
Многие печеночники — слоевищные (талломные) организмы. Типичным представителем слоевищных печеночников является маршанция многообразная.
Однако большинство печеночников не талломные, а облиствленные. Листья у печеночников, в отличие от листьев листостебельных мхов, расположены не но спирали, а 3-4 рядами.
Ризоиды одноклеточные. Протонема у большинства печеночников развита слабо и недолговечна.
Обитают печеночники на влажной почве, на скалах, по берегам рек.
Очень хорошо развито у печеночников вегетативное размножение.
Класс листостебельные, или настоящие мхи
Класс листостебельные, или настоящие мхи (Bryopsida, или Musci) — это самый крупный класс моховидных, насчитывающий около 25 000 видов.
Класс листостебельные включает три подкласса:
- бриевые;
- сфагновые;
- андреевые мхи.
Бриевые мхи
Подкласс бриевые (Bryidae), или зеленые мхи, включает 14 000 видов. В сырых местах представители этой группы распространены повсеместно: они поселяются на почве, коре, стволах деревьев. Зеленые мхи обитают в еловых и сосновых лесах, на болотах, на склонах гор, а в тундре часто образуют сплошной покров.
Типичный представитель зеленых мхов — кукушкин лен, или политрихум (Polytrichum commune) — Бриевые мхи семейств политриховые и даусониевые — единственные представители моховидных, гаметофиты которых имеют относительно хорошо развитые проводящие ткани, напоминающие ксилему и флоэму примитивных сосудистых растений. Листья на гаметофитах всех бриидов расположены но спирали. Верхняя сторона листа покрыта столбцами фотосинтезирующих клеток ассимиляционными пластинками. Покровная ткань (эпидерма), защищающая растение от высыхания, расположена только на нижней стороне листа. Стебель и листья политриховых содержат также механические ткани, представляющие собой удлиненные клетки, напоминающие склереиды сосудистых растений. Мхи из семейства политриховые — многолетние, относительно крупные растения (например, высота стеблей кукушкина льна иногда достигает 40-50 см), часто образующие обширные покрытия на почве в лесах, на болотах и в тундре.
Гаметофит всех зеленых мхов представляет собой облиствленный стебель, а спорофит — коробочку на ножке. На ранней стадии развития спорофит зеленый и способен к фотосинтезу. По мере созревания он желтеет, затем становится оранжевым и, наконец, бурым, утрачивает способность к фотосинтезу и полностью переходит к питанию за счет гаметофита (паразитирует на гаметофите).
Ризоиды зеленых мхов, в отличие от ризоидов печеночников, — многоклеточные, но всасывают воду относительно плохо. Поэтому зеленые мхи, так же как и другие классы мхов, впитывают воду всей поверхностью тела, в первую очередь листьями. Так, кукушкин лен может впитать воды в 4-5 раз больше сухой массы собственного тела. В связи с этим мхи часто заболачивают почву, на которой растут.
Характерной особенностью жизненного цикла зеленых мхов является то, что их развитие начинается с образования особой нитчатой структуры — протонемы, внешне напоминающей нитчатую зеленую водоросль. Интересно, что у некоторых листостебельных мхов гаметофит вообще не развивается. Главной жизненной формой таких мхов становится протонема. Наиболее известен из них светящийся мох схистостега перистая (Shistostega реnnаtа), обитающий в пещерах но всей южной Европе. Именно с ним связано появление легенд о сокровищах гномов, исчезающих с рассветом.
У зеленых мхов, так же как у печеночников, хорошо развито вегетативное размножение.
Сфагновые мхи
Подкласс сфагновые (Sphegnidae), или белые мхи, представлен единственным родом сфагнум (Sphagnum), включающим свыше 300 видов. Характерной особенностью сфагнума является ветвящийся стебель: от главного стебля у сфагнума отходят не отдельные листья, а мутовки ветвей (иногда но 5 в узле), а на верхушке побега образуется головка из сближенных веточек.
Начальной фазой развития сфагнума является образование из споры пластинчатой протонемы.
Сфагнум не имеет ризоидов. По мере роста растения нижние части стебля отмирают и погружаются на дно. В процессе роста сфагнум не только заболачивает почву, но и подкисляет воду до pH ниже 4. В кислой среде без доступа кислорода отмершие стебли сфагнума и других растений не сгнивают, а превращаются в торф.
Торфяные болота — интересный объект для археологов и палеоботаников. В кислой среде торфяных болот прекрасно сохраняются споры древних растений, стволы деревьев, древние орудия труда, лодки, строительные сооружения. Так, недавно на территории Великобритании в торфяных отложениях была обнаружена деревянная дорога, соединявшая два поселения людей каменного века. Возраст этого сооружения — 6000 лет.

Рис. 2. Мох сфагнум: а) общий вид; 6) коробочка; в) клетки листа под микроскопом
Торф — прекрасное, причем возобновляемое топливо. В основном торф используют в тепловых электростанциях для получения электроэнергии. В сельском хозяйстве торф применяют в качестве удобрения, а также для удержания влаги в почве. В тепличном хозяйстве торфо-перегнойные горшочки служат для выращивания рассады.
В медицине сфагнум используют как прекрасный перевязочный материал и наполнитель для различных поглощающих влагу прокладок. Сфагнум по сравнению с обычными перевязочными материалами, например вагой, впитывает влагу в 5-6 раз эффективнее. Кроме того, в отличие от ваты, сфагнум обладает выраженными бактерицидными свойствами.
Интересной особенностью сфагнума является механизм рассеивания спор.
Спорофит сфагнума представляет собой круглую коробочку, возвышающуюся на подставке (ложноножке) из тканей гаметофита. Во влажную погоду через устьица в коробочку проникает воздух. При подсыхании коробочки устьица на се поверхности закрываются, давление воздуха внутри повышается, в результате с отчетливым хлопком крышечка срывается и над коробочкой поднимается облачко спор.
Торфяные болота занимают около 1% площади Земного шара и играют важную роль в регулировании водного баланса Земли. Запас воды из верховых болот питает реки.
Андреевые мхи
Подкласс андреевые (Andreaeidae), или черные мхи, объединяет около 120 видов черно-зеленых или красно-бурых наскальных мхов, характерных для горных и арктических областей. Протонема — пластинчатая, толстостенная, многолопастная.
Интересен механизм рассеивания спор. Коробочка на ложноножке из тканей гаметофита растрескивается на 4 пластинки. В сухую погоду за счет сокращения центральной оси она, как елочная игрушка, сжимается, и споры через открывшиеся щели высыпаются из коробочки. Во влажную погоду ось коробочки удлиняется и щели закрываются.
Отдел моховидные – это высшие споровые растения, видовое разнообразие которых достигает 20 тыс. Изучение мхов ведется много столетий, ученых занятых их исследованием прозвали бриологами, они основали отдельную ботаническую отрасль, посвященную мохообразным – бриологию. Бриология – наука о мхах, изучает строение, размножение и развитие моховидных (собственно мхов, печеночников, антоцеротовых).
Общая характеристика мхов

Моховидные – одни из древнейших растений, населяющих нашу планету. Остатки найдены в окаменелостях конца палеозойской эры. Распространение мхов связано с предпочтением влажной среды и затенённой местности, поэтому большинство населяют северную часть Земли. Плохо приживаются на засоленных территориях и пустынях.
Классы моховидных
Листостебельные мхи – самый многочисленный класс. Растения состоят из стебля, листьев и ризоидов.
Стебель может расти вертикально или горизонтально, разделен на кору и основную ткань (содержит воду, крахмал, хлоропласты для фотосинтеза).
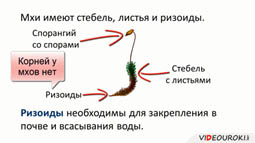
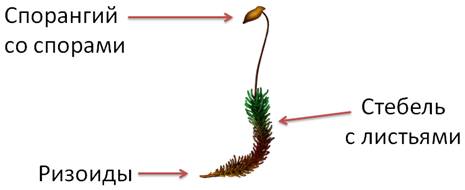
Клетки стебля могут давать нитевидные отростки – ризоиды, необходимы для крепления к почве и поглощения воды. Они чаще находятся у основания стебля, но могут покрывать его по всей длине.
Листья простые, часто крепятся к стеблю под прямым углом, по спирали. Листовые пластинки оснащены хлоропластами, в центре располагается жилка (служит для проведения питательных веществ).
Лиственные мхи могут размножаться стеблями, почками, ветвями, которые дают побег, так формируются сплошные ковры из мхов, устилающие землю. К классу листостебельных относятся сфагновые (имеют разнообразный окрас стебля — светло-зелёный, желтый, красный), андреевые и бриевые мхи.

Сфагновый мох
Печеночники встречаются на побережьях, болотах, скалистой местности. Отличительные черты: листья не имеют жилки, дорсовентральное строение, особый механизм раскрытия спорофита.
Листья расположены рядами, имеют две лопасти (нижняя лопасть, часто, завёрнутая и служит как резервуар для воды), ризоидные отростки – одноклеточные. Во время высыпания спор, спорофитовая коробочка раскрывается на отдельные створки, а рассеиванию клеток способствуют элатеры (пружинные образования).
Размножение может осуществляться с помощью почек (вегетативно), которые образуются на верхнем полюсе листьев. Представители класса пеллия эндиевлистная, милия аномальная, мох маршанция и др.
Антоцеротовые мхи населяют тропическую зону. Многоядерное тело (таллом) имеет розетковидную форму, состоит из однотипных клеток. В верхних шарах клеток находятся хроматофоры (содержат темно-зеленый пигмент). Нижняя часть таллома дает отростки, ризоиды, само тело образует полости, заполненные вязкой жидкостью, которая поддерживает постоянную влагу.
На поверхности таллома при неблагоприятных условиях образуются клубни, устойчивые к низкой влажности, после периода засухи формируют новое поколение. Растения однодомные, органы размножения развиваются в толще таллома, стадия спорофита преобладающая. К антоцеротовым относятся фолиоцерос, антоцерос, нототилас и др.

Как размножаются мхи?
Идет чередование бесполого и полового способа размножение в жизненном цикле мхов. Бесполый период начинается с образования спор и прорастания их на влажной почве (формируется предросток, тонкая нить, которая дает жизнь мужским и женским особям). Существует два вида мхов:
Однодомные – мужские и женские органы размножения находятся на одном растении.
Двудомные – органы размножения находятся у разных представителей пола.
После прорастания споры, жизненный цикл мха переходит в половую фазу. Органы полового размножения – антеридии (мужские) и архегонии (женские). Представители мужских особей слабее женских, меньших размеров, после формирования антеридиев отмирают.

Процесс размножения мхов
Сперматозоиды образуются на мужских растениях, яйцеклетки – на женских, после их слияния формируется зигота (находится на женской особи, она питает незрелый спорофит), которая в дальнейшем развивается в спорангий. После созревания спорангия, он раскрывается, из него высыпаются споры – начинается снова бесполый период размножения мхов.
Воспроизведение потомства возможно вегетативным способом, мхи образуют талломы (зеленые ответвления), почки, клубни, которые на влажной почве хорошо приживаются.
Каково значение спор в жизни мхов?
Споры – это клетки, необходимые для размножения мхов. Моховые растения не цветут, не имеют корней, поэтому для продолжения рода у них сформировался спорофит со спорангиями (место созревания спор).
Спорофит имеет короткий жизненный цикл, после высыхания споры рассеиваются вокруг, при попадании на влажную почву быстро приживаются. Могут при неблагоприятных условиях долго сохраняться, не прорастая, устойчивые к низким и высоким температурам, длительным засухам.
Значение мхов в природе и жизни человека
Мхи – это пища для многих беспозвоночных животных.
После отмирания дают залежи торфа, который необходим в производстве пластмасс, смол, карболовой кислоты, используется как топливо или удобрение.
Мох полностью укрывает землю в местах роста, что приводит к накоплению влаги и заболачиванию территории. Таким образом, прорастание другой растительности становится невозможным. В то же время они предотвращают эрозию, разрушение почв поверхностными водами и ветрами. Когда мхи отмирают, берут участие в образовании почвы.
Способны расти на местах пожаров, стойкие и выносливые, они населяют территорию тундры (основной растительный фон, так как другие растения не могут выжить в таких условиях).
В военное время сфагновый мох использовали, как перевязочный материал из-за его бактерицидных свойств, и способности впитывать влагу.
С помощью мхов можно ориентироваться на местности: они не любят света, поэтому располагаются с теневой стороны камней, деревьев. Мох указывает человеку на север.

Видеоурок знакомит учащихся со споровыми растениями. Рассматриваются особенности строения печёночных, зелёных и сфагновых мхов. В уроке представлена классификация мхов. Множество иллюстраций поможет учащимся лучше усвоить знания о данной группе живых организмов.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Мхи. Многообразие. Строение мхов, их значение"
В отличие от водорослей мхи имеют стебель и листья, за исключением ряда видов примитивных печёночных мхов, у которых тело представлено слоевищем. Настоящих корней у мхов нет, их заменяют ризоиды, которыми они укрепляются в почве и всасывают воду.
Так как тело мхов расчленено на стебель и листья, а размножаются они спорами, то их относят к высшим споровым растениям.
Мхи — это многочисленная обособленная и очень древняя группа высших растений. На нашей планете насчитывается около 28 тыс. видов мхов. Это небольшие растения. Их высота колеблется от нескольких миллиметров до нескольких сантиметров. Очень редко можно встретить мхи более 20 см высотой. Мхи распространены преимущественно в хорошо увлажнённых местах. Их можно встретить на почве в лесах, пустынях и на болотах, на камнях и стволах деревьев, на стенах и крышах старых домов, в воде пресных водоёмов. Не встречаются мхи только в морях и на засоленных почвах. Многие виды мхов способны переносить неблагоприятные условия в покоящемся состоянии. При наступлении благоприятных условий такие мхи переходят в активное состояние и продолжают свой рост.
Мхи устроены проще, чем остальные высшие растения. У них имеются фотосинтезирующие, а также покровные и механические ткани. У наиболее сложно устроенных мхов появляются и специальные клетки, выполняющие проводящую функцию. Но типичные проводящие ткани у мхов отсутствуют. Появление у мхов тканей — это результат их приспособления к наземному образу жизни.
Различают печёночные и листостебельные мхи.
Представителями печёночных мхов являются риччия и маршанция. Те, у кого дома есть аквариум, хорошо знают плавающее растение, зелёным ковром покрывающее поверхность воды. Это один из печёночных мхов — риччия. Тело её состоит из вильчато разветвлённого слоевища. При хорошем освещении риччия быстро разрастается, образуя плотные подушки на поверхности воды.

Маршанция — это многолетнее растение, имеющее вид зелёного лопастного листка, стелющегося по поверхности земли.

Один из самых известных листостебельных зелёных мхов — кукушкин лён, его часто можно встретить в заболоченных или просто во влажных местах. Это многолетнее растение, высотой до 20 см. Его прямостоячий стебель несёт узкие мелкие листья.

У кукушкина льна есть мужские и женские растения. На верхушках мужских растений размещаются половые органы, в которых развиваются подвижные половые клетки (гаметы) — сперматозоиды.
У женских растений на верхушках расположены половые органы с женской половой клеткой — яйцеклеткой.
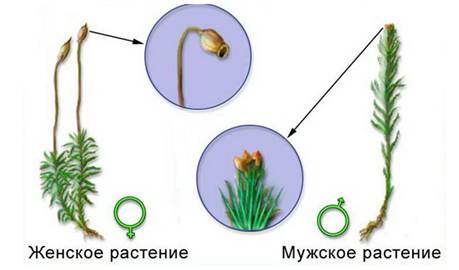
При наличии капелек влаги гаметы сливаются и образуют зиготу.
На женских растениях развиваются коробочки на длинных ножках, покрытые волосистыми заострёнными колпачками. Они напоминают сидящую кукушку. Отсюда и название мха — кукушкин лён. В коробочках развиваются споры. Зрелые споры высыпаются из коробочки и образуют зелёную нить. Из почек этой нити вырастают листостебельные растения мха.
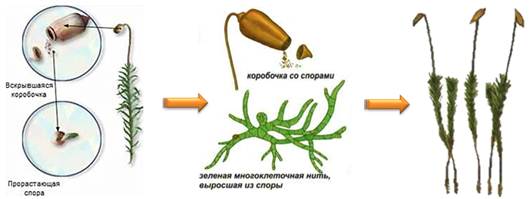
Представителем белых, или сфагновых, мхов является сфагнум.
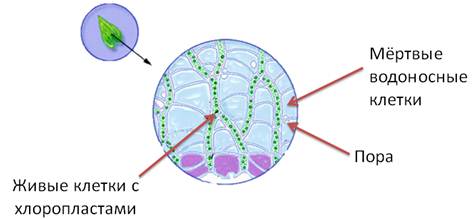
Сфагнум — растение с сильно ветвящимся стеблем. В отличие от других зелёных мхов он не имеет ризоидов. Стебель и ветви большинства видов сфагнума покрыты мелкими светло-зелёными листьями.

Каждый лист состоит из клеток двух типов. Живые, узкие, сильно вытянутые в длину клетки, в которых содержатся хлоропласты, расположены между очень крупными мёртвыми водоносными клетками. В клеточных стенках водоносных клеток есть поры. Через эти поры внутрь клеток поступает вода, которая может в них удерживаться. Из-за большого количества водоносных клеток в листьях и стебле сфагновый мох может быстро впитывать большое количество воды (в 20 — 40 раз больше своей сухой массы).
Размножается сфагнум спорами, так же как кукушкин лён и другие мхи. На концах верхних ветвей у него образуются маленькие коробочки, в которых созревают споры.

Обычно сфагнум растёт на верховых болотах, покрывая поверхность почвы сплошным ковром, но он может расти и под пологом леса среди кукушкина льна. Там, где поселился сфагнум, почвы переувлажнены. На избыточно влажной почве деревья растут плохо, становятся угнетёнными, а сфагнум, напротив, разрастается пышным ковром, и лес постепенно заболачивается. Сфагновый мох растёт верхней частью побегов, нижние их части отмирают. Мох образует кислоты, которые тормозят гниение отмирающих стеблей. Медленное разложение этих стеблей во влажной среде при небольшом количестве кислорода приводит к образованию полезного ископаемого – торфа, который широко используется человеком. Торф применяется как удобрение, топливо и сырьё для промышленности.
Значение мхов в природе велико. Зачастую мхи, так же как и лишайники, первыми покрывают голые скалы и другие участки, лишённые растительного покрова. Своими ризоидами они постепенно разрушают горные породы, в результате чего образуется почва, на которой могут поселиться другие растения.
Мхи играют также очень важную роль в регулировании водного режима планеты. Мхи впитывают и удерживают большое количество воды. Болота, на которых часто преобладают сфагновые мхи, нередко дают начало ручьям и рекам. Поэтому осушение больших площадей болот может приводить к обмелению рек, изменению климата, почв, обеднению растительного и животного мира. С другой стороны, развиваясь плотным ковром на используемых человеком землях, мхи затрудняют дыхание почвы и вызывают её заболачивание. Сфагновые мхи содержат бактерицидные вещества и поэтому могут быть использованы в медицине.
Читайте также: