
Пример проявления аллельных генов это урожайность и процент белка
Добавил пользователь Алексей Ф. Обновлено: 18.09.2024
В генотип каждого живого организма входит огромное количество разнообразных генов. Аллельные гены – одни из них. Наравне с другими они образуют органическую совокупность, оказывают влияние на реализацию общих функций.
Что такое аллельные гены
Аллельными называются гены, занимающие идентичное местоположение в гомологичных хромосомах и отвечающие за развитие одного признака.
Наличие в диплоидном организме двух различных аллелей одного гена приводит к гетерозиготности. И, наоборот, присутствие ничем не отличающихся – признак гомозиготного организма.
Осторожно! Если преподаватель обнаружит плагиат в работе, не избежать крупных проблем (вплоть до отчисления). Если нет возможности написать самому, закажите тут.
Главное отличие аллельных генов от неаллельных заключается в их расположении. Неаллельные имеют привычку располагаться в разных местах. Кроме того, они призваны кодировать не один и тот же признак, а разные.
Где расположены гены
Все гены расположены в хромосомах – миниатюрных элементах, хранящих определяющий объем наследственной информации. В свою очередь, хромосомы привыкли находиться в ядре – самом крупном органоиде клетки.
Способы взаимодействия аллельных генов
Находясь в одном организме, аллели вынуждены взаимодействовать между собой. В большинстве случаев один из них подавляет проявление другого. Тогда говорят о том, что первый является доминантным, а второй – рецессивным. А сам процесс называют полным доминированием.
Однако за время изучения вопроса учеными было зафиксировано несколько разных видов взаимодействия. Несмотря на то, что они встречаются реже, каждый из них играет важную роль в реализации генетической информации, обуславливает необходимую для эволюции многовариантность.
Кодоминирование
Кодоминирование подразумевает полное отсутствие рецессивно-доминантных отношений. Это тот случай, когда оба аллеля проявляют себя в полной мере и никак не взаимодействуют друг с другом.
Простым примером является наследование человеком той или иной группы крови. Результатом кодоминирования здесь является получение гибридом нового, отличающегося от родительских признака, а не их усредненного варианта.
Сверхдоминирование
Сверхдоминированием называется такой тип взаимодействия, при котором фиксируется более яркое проявление подконтрольного признака у гетерозиготного организма, чем у любого гомозиготного.
Генетики считают, что именно он стал причиной формирования такого явления как гетерозис. Когда дочерний организм значительно превосходит родительские по плодовитости, жизнеспособности, продуктивности, выносливости, темпам роста.
Полное и неполное доминирование
При полном доминировании получившийся в результате скрещивания фенотип демонстрирует исключительно признак доминантного аллеля. В то время как неполное доминирование предполагает неполное подавление рецессивной формы.
Скрещивание цветка с красными лепестками и полностью белого цветка в результате дает получение однотонных розовых гибридов. При этом при последующем скрещивании гибридов можно получить красные, розовые и белые цветы.
За что отвечают аллельные гены
Обнаружить аллельные гены можно в организмах людей, рыб, животных, насекомых, грибов, растений. Они отвечают за передачу разнообразных наследственных характеристик. Например:
- цвета глаз;
- оттенка кожи;
- длины шерстного покрова;
- густоты меха;
- группы крови;
- окраски семян;
- формы рисунка на лепестках.
Зафиксированы ситуации, при которых аллельные гены отвечают за жизнь своего носителя. Такие аллели называются летальными и возникают из-за произошедшей мутации. Они могут проявлять себя в постэмбриональном и эмбриональном периодах, всегда ведут к развитию несовместимых с жизнью пороков. Печальным примером может служить отличающийся мутантным доминантным аллелем синдром Гентингтона.
Примеры взаимодействия аллельных генов в природе
Первым и самым известным примером взаимодействия аллельных генов в природе является результат опыта известного австрийского биолога, основоположника учения о наследственности Грегора Иоганна Менделя.
В ходе работы над гибридизацией гороха ученый понял, что скрещивание растений с зелеными и желтыми горошинами приводит к выведению сорта исключительно с желтыми горошинами. Так была выведена первая теория о полном доминировании одного из алеллей.
Примеры неполного доминирования
Образцы влияния неполного доминирования на формирование конкретного признака можно наблюдать при наследовании окраски растениями: усредненные оттенки ягод, цветов.
Яркий пример – получение пшеницы с укороченными стеблями при скрещивании карликовых пород и растений с длинными стеблями.
В мире животных та же ситуация наблюдается при разведении куриц: скрещивание черных и белых особей приводит к получению однотонного серебристого потомства.
Примеры сверхдоминирования
К примерам сверхдоминирования могут относиться ситуации с биохимическими особенностями особей.
У популяции людей, проживающих в регионах с неблагоприятной эпидемиологической обстановкой по малярии, повышена частота мутантного аллеля-защитника. Имея его, гетерозиготный организм легко справляется с болезнью, находится в состоянии повышенной жизнеспособности.
У животных образцами сверхдоминирования могут считаться лучшая приспособленность к внешней среде, нежели у гомозиготных предков, повышенная выносливость, плодовитость.
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Ген является структурной единицей наследственной информации. Материально ген представлен участком молекулы ДНК (в редких случаях – РНК). Гены контролируют элементарные признаки в процессе индивидуального развития организма. Первые исследования природы гена, проводившиеся в начале XX в., в основном были направлены на выяснение роли гена в передаче наследственных признаков. Не менее важной задачей стала расшифровка закономерностей действия генов. Ее решение имеет не только теоретическое, но и практическое значение, поскольку позволит предотвратить возможные вредные последствия этого действия.
Исследования генетиков установили дискретный характер генов, что подтверждается их независимым друг от друга наследованием: каждый из генов определяет развитие какого–то признака, независимого от других. Однако между различными генами существуют разные типы взаимодействия, обусловленные сложными отношениями как между аллельными, так и между неаллельными генами. Объединяясь в генотипе, они все вместе выступают как система взаимодействующих между собой генов особи.
Среди взаимодействий генов следует назвать прежде всего отношения доминантности и рецессивности, когда рецессивная аллель гена под влиянием доминантной аллели не проявляется в фенотипе. Кроме того, имеются факты, показывающие, что гены влияют на проявление действий других, неаллельных генов. Описаны также случаи, когда развитие того или иного признака организма находится под контролем не одного, а многих генов. Например, у человека не менее четырех генов определяют различие в цвете кожи представителей негроидной и европеоидной рас.
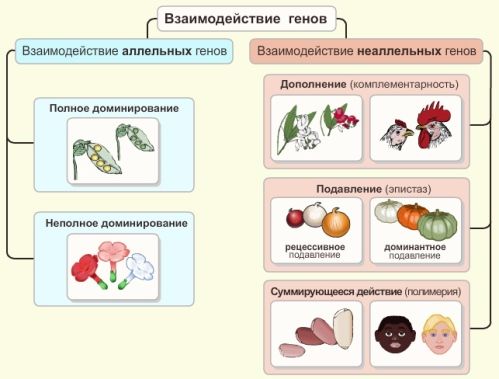
Взаимодействие генов
| Взаимодействие аллельных генов | Полное доминирование – одна аллель гена (рецессивная) под влиянием доминантной аллели не проявляется в фенотипе |
| Неполное доминирование наблюдается промежуточный характер наследования признаков у гетерозиготной особи с большим или меньшим отклонением в сторону доминантного или рецессивного состояния | |
| Кодоминирование – совместное участие обоих аллелей в определении признака у гетерозиготной особи | |
| Взаимодействие неаллельных генов | Комплементарное взаимодействие – доминантные или рецессивные аллели обусловливают развитие нового, отличного от родительских вариантов, признака |
| Множественное действие генов – действие одного гена влияет на развитие нескольких признаков | |
| Полимерия – степень развития одного и того же признака обусловлена влиянием целого ряда генов | |
| Эпистаз – аллели одного гена подавляют проявление аллелей других генов |
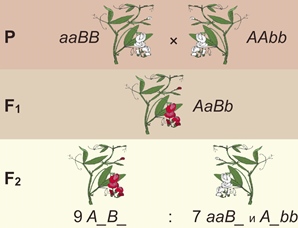
Комплемертансть. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента, вызывающего окраску цветка. Ген В определяет синтез фермента, под действием которого образуется пигмент. Поэтому, например, цветки душистого горошка с генотипом ааВB и АAbb имеют белый цвет: в первом случае вырабатывается фермент, но не вырабатывается вещество, на которое он действует; во втором — есть предшественник пигмента, но нет фермента, переводящего его в пурпурный пигмент.
При скрещивании двух таких растений с белыми цветками в первом поколении все растения имеют пурпурный венчик, а во втором поколении наблюдается расщепление 9 (пурпурные цветки) : 7 (белые цветки).
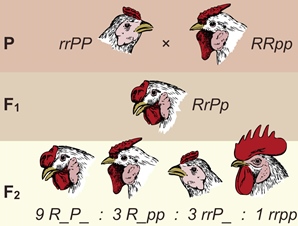
При скрещивании петуха с розовидным гребнем с курицей, имеющей гороховидный гребень, в F1 следует ожидать доминирования одного из признаков, а в F2 — расщепления 3:1.
В действительности результаты скрещивания оказываются резко отличными от ожидаемых. В F 1 все птицы имеют гребни, не похожие ни на одного из родителей, — эти гребни называются ореховидными. В F 2 , полученном от скрещивания петухов и кур с ореховидными гребнями, обнаруживаются четыре фенотипических класса: два с проявлением родительских признаков (розовидный и гороховидный гребни) — по 3 части от всего потомства, 9 частей с ореховидным гребнем и 1 часть с новым типом гребня (простым, или листовидным).
Наблюдаемое расщепление по фенотипу 9:3:3:1 ясно указывает, что имеет место дигибридное скрещивание. Для объяснения необычного характера наследования формы гребня была предложена гипотеза, согласно которой неаллельные гены R и Р , определяющие развитие гребня определенной формы, взаимодействуют друг с другом, причем результаты взаимодействия зависят от того, в какой форме находится каждый из генов — в доминантной или в рецессивной.
R — розовидный гребень, r — отсутствие розовидного гребня, P — гороховидный гребень, p — отсутствие гороховидного гребня. Если в обеих парах генов присутствуют доминантные аллели R_P_, то возникает новая форма гребня — ореховидный гребень, при генотипе rrpp у кур развивается листовидный гребень.
Комплементарным называется взаимодействие, при котором действие генов из одной пары дополняется действием генов из другой пары таким образом, что в результате появляется новый признак.
Отличительная особенность комплементарного взаимодействия от закона независимого наследования признаков заключается в количестве учтённых признаков: в третьем законе рассматривается комбинирование двух признаков, а в комплементарном взаимодействии четыре вариации одного признака.
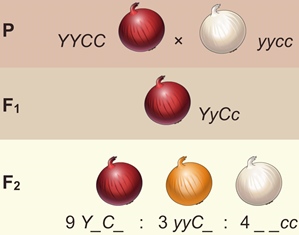
Рецессивный эпистаз. Скрещивание лука с красными и белыми луковицами даёт в F 1 растения только с красными луковицами, а в F 2 — расщепление: 9/16 — с красными, 3/16 — с жёлтыми и 4/16 — с белыми луковицами.
Обозначим ген, определяющий красный цвет луковиц, через Y ; ген, определяющий жёлтый цвет, — через y . Рецессивный ген с (от англ. colorless — бесцветный) в гомозиготном состоянии определяет образование белых луковиц, а доминантный С — окрашенных.
Взаимодействие генов, при котором один ген подавляет действие другого, называется эпистазом (от греч. эпи — над и стазис — стоять).
Если эпистатический ген проявляется в рецессивной форме, то такое взаимодействие называют рецессивным эпистазом. Расщепление по фенотипу при такой форме взаимодействия генов 9:3:4.
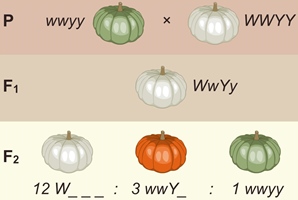
Доминантный эпистаз. В некоторых случаях наличие одного доминантного гена подавляет проявление другого неаллельного гена. Например, ген Y обуславливает жёлтый цвет плодов тыквы, а его рецессивная аллель y — зелёный цвет плодов.
При наличии в генотипе доминантной аллели особого гена W действие гена Y (или y) не проявляется и плоды имеют серовато-белую окраску. При генотипе wwY_ плоды тыквы имеют жёлтый цвет. Наконец, если оба гена представлены рецессивными аллелями ( wwyy ), плоды тыквы имеют зелёный цвет. При скрещивании тыкв с белыми ( WWYY ) и зелёными ( wwyy ) плодами все гибриды первого поколения будут иметь белые плоды ( WwYy ), а во втором поколении наблюдается расщепление по фенотипу 12 (белая кожура): 2 (жёлтая кожура): 1 (зелёная кожура).
Если эпистатический ген проявляется в доминантной форме, то такое взаимодействие называют доминантным эпистазом.
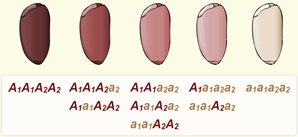
Полимерия. У пшеницы красный цвет зёрен определяется двумя генами: А1 и А2. Неаллельные гены обозначаются одной буквой алфавита потому, что определяют развитие одного признака. При генотипе А1А1А2А2 окраска зёрен наиболее интенсивная. При генотипе а1а1а2а2 зёрна имеют белый цвет. В зависимости от числа доминантных генов в генотипе можно получить все переходы между интенсивно красной и белой окраской.
Такая обусловленность определённого признака несколькими парами неаллельных генов, обладающих одинаковым действием, носит название полимерии (от греч. поли — много и мерос — часть).
При кумулятивной (от лат. кумуло — собираю, накапливаю), или накопительной, полимерии, как в случае с окраской зёрен пшеницы, степень проявления признака зависит от суммирующего проявления генов.

У человека интенсивность окраски кожи определяется несколькими парами генов. Учёные предполагают, что цвет кожи зависит от трёх пар генов. Самая тёмная кожа (у представителей негроидной расы) будет определяться генотипом А1А1А2А2А3А3; у мулатов будут присутствовать несколько рецессивных аллелей, а у коренного населения Европы генотип не будет иметь доминантных генов — а1а1а2а2а3а3.
В зависимости от числа доминантных генов в генотипе может формироваться более светлый или более тёмный цвет кожи.
С точки зрения генетики, наиболее изученной является система АВ0, определяющая I (0), II (А), III (В) и IV (АВ) группы крови. На поверхности эритроцитов могут находиться агглютиногены (антигены) А и В, а в плазме крови – агглютинины (антитела) и . В норме одноименные агглютиногены и агглютинины совместно не обнаруживаются. Нужно отметить, что А- и В-антигены образуют многочисленный ряд антигенов (А1, А2 . A; В1, В2 …В).
Понятие о комплиментарном взаимодействии генов. Примеры.
Эпистатическое взаимодействие генов. Примеры.
Подавляющий ген – эпистатический
Подавляемый ген – гипостатический
Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого различают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При эпистатичном взаимодействии генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.
Если ген-супрессор рецессивный, то возникает криптомерия (греч. хриштад - тайный, скрытый).
У человека таким примером может быть "Бомбейский феномен". В этом случае редкий рецессивный аллель "h" в гомозиготном состоянии (hh) подавляет активность гена jB (определяющий В (III) группу крови системы АВО). Поэтому женщина с генотипом jв_hh, фенотипно имеет I группу крови - 0 (I).
При эпистазе один из генов (В) выражается фенотипически лишь при отсутствии в генотипе определенного аллеля другого гена (А). В его присутствии действие гена В не проявляется. В строгом смысле слова, этот вид взаимодействия неаллельных генов может быть рассмотрен как вариант взаимодополняющего действия определенных аллелей этих генов, когда один из них способен обеспечить развитие признака, но лишь в присутствии определенного аллеля другого гена. В этой ситуации фенотип организма зависит от конкретного сочетания аллелей неаллельных генов в их генотипах и расщепление по фенотипу в потомстве двух дигетерозигот по этим генам может быть различным.
При доминантном эпистазе, когда доминантный аллель одного гена (А) препятствует проявлению аллелей другого гена (В или b), расщепление в потомстве зависит от их фенотипического значения и может выражаться соотношениями 12:3:1 или 13:3 (рис. 6.19). При рецессивном эпистазе ген, определяющий какой-то признак (В), не проявляется у гомозигот по рецессивному аллелю другого гена (аа). Расщепление в потомстве двух дигетерозигот по таким генам будет соответствовать соотношению 9:3:4 (рис. 6.20). Невозможность формирования признака при рецессивном эпистазе расценивают также как проявление несостоявшегося комплементарного взаимодействия, которое возникает между доминантным аллелем эпистатического гена и аллелями гена, определяющего этот признак.
Полимерия и ее роль в детерминации количественных признаков. Аддитивный эффект.
Важная особенность полимерии - суммация действия неаллельних генов на развитие количественных признаков. Если при моногенном наследовании признака возможны три варианта "доз" гена в генотипе: АА, Аа, аа, то при полигенных количество их возрастает до четырех и более. Суммация "доз" полимерных генов обесчивает существования непрерывных рядов количественных изменений.
Биологическое значение полимерии заключается еще и в том, что признаки, кодируемые этими генами, более стабильны, чем те, которые кодируются одним геном. Организм без полимерных генов был бы очень неустойчивым: любая мутация или рекомбинация приводила бы к резкой изменчивости, а это в большинстве случаев имеет неблагоприятный характер.
У животных и растений есть много полигенных признаков, среди них и ценные для хозяйства: интенсивность роста, скороспелость, яйценоскость, количество молока, содержание сахаристых веществ и витаминов и т.п.
Пигментация кожи у человека определяется пятью или шестью полимерными генами. В коренных жителей Африки (негроидной расы) преобладают доминантные аллели, у представителей европеоидной расы - рецессивные. Поэтому мулаты имеют промежуточную пигментацию, но при браках мулатов у них возможно появление как более, так и менее интенсивно пигментированных детей.
Многие морфологические, физиологические и патологические особенности человека определяются полимерными генами: рост, масса тела, величина артериального давления и др. Развитие таких признаков у человека подчиняется общим законам полигенного наследования и зависит от условий среды. В этих в случаях наблюдается, например, склонность к гипертонической болезни, ожирению и др. Данные признаки при благоприятных условиях среды могут не проявиться или проявиться незначительно. Эти полигенные признаки отличаются от моногенных. Изменяя условия среды можно обеспечить профилактику ряда полигенных заболеваний.
Наследование признаков при полимерном взаимодействии генов. В том случае, когда сложный признак определяется несколькими парами генов в генотипе и их взаимодействие сводится к накоплению эффекта действия определенных аллелей этих генов, в потомстве гетерозигот наблюдается разная степень выраженности признака, зависящая от суммарной дозы соответствующих аллелей. Например, степень пигментации кожи у человека, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4). При браке двух мулатов, гетерозиготных по всем четырем парам, которые образуют по 2 4 = 16 типов гамет, получается потомство, 1/256 которого имеет максимальную пигментацию кожи, 1/256 — минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака. В разобранном примере доминантные аллели полигенов определяют синтез пигмента, а рецессивные — практически не обеспечивают этого признака. В клетках кожи организмов, гомозиготных по рецессивным аллелям всех генов, содержится минимальное количество гранул пигмента.
Плейотропное действие генов. Примеры.
Назовите основные биометрические характеристики, испльзуемые при генетико-математическом анализе количественных признаков.
Физиологические — относятся к форме тела. В качестве примера можно привести: отпечатки пальцев, распознавание лица, ДНК, ладонь руки, сетчатка глаза, запах, голос.
Поведенческие — связаны с поведением человека. Например, походка и речь. Порой используется термин англ. behaviometrics для этого класса биометрии.
Понятие о варианте и вариационном ряде.
Основные обозначения вариационного ряда
р — частота ("вес") варианты, число ее повторений в вариационном ряду;
n — общее число наблюдений (т.е. сумма всех частот, n = Σр);
Vmax и Vmin — крайние варианты, ограничивающие вариационный ряд (лимиты ряда);
А — амплитуда ряда (т.е. разность между максимальной и минимальной вариантами,А = Vmax — Vmin)
Виды вариаций:
а) простой — это ряд, в котором каждая вариата встречается по одному разу (р=1);
6) взвешенный — ряд, в котором отдельные варианты встречаются неоднократно (с разной частотой).
Сущность средней арифметической, среднего квадратического отклонения, дисперсии и методы их расчета.
для оценки состояния здоровья — например, параметров физического развития (средний рост, средняя масса тела, среднее значение жизненной емкости легких и др.), соматических показателей (средний уровень сахара в крови, средняя величина пульса, средняя СОЭ и др.);
для оценки организации работы лечебно-профилактических и санитарно-противоэпидемических учреждений, а также деятельности отдельных врачей и других медицинских работников (средняя длительность пребывания больного на койке, среднее число посещений на 1 ч приема в поликлинике и др.);
для оценки состояния окружающей среды.
Суммировать варианты: V1+V2+V3+. +Vn = Σ V;
Сумму вариант разделить на общее число наблюдений: М = Σ V / n
Получить произведение каждой варианты на ее частоту — Vp
Найти сумму произведений вариант на частоты: V1p1 + V2p2+ V3p3 +. + Vnpn = Σ Vp
Полученную сумму разделить на общее число наблюдений: М = Σ Vp / n
Среднее квадратичное отклонение определяется как обобщающая характеристика размеров вариации признака в совокупности. Оно равно квадратному корню из среднего квадрата отклонений отдельных значений признака от средней арифметической, т.е. корень из дисперсии и может быть найдена так:
1. Для первичного ряда:

2. Для вариационного ряда:

Преобразование формулы среднего квадратичного отклонени приводит ее к виду, более удобному для практических расчетов:

Среднее квадратичное отклонение определяет на сколько в среднем отклоняются конкретные варианты от их среднего значения, и к тому же является абсолютной мерой колеблемости признака и выражается в тех же единицах, что и варианты, и поэтому хорошо интерпретируется.
Найти отклонение (разность) каждой варианты от среднеарифметической величины ряда (d = V — М);
Возвести каждое из этих отклонений в квадрат (d2);
Получить произведение квадрата каждого отклонения на частоту (d2р);
Найти сумму этих отклонений: d21p1 + d22p2 + d23p3 +. + d2npn = Σ d2р;
Полученную сумму разделить на общее число наблюдений (при n Фенотипическая дисперсия - статистическая величина, с помощью которой оценивается размах вариативности какого-либо количественного признака в популяции.

На этом уроке подробно рассмотрены примеры решения задач на аллельное и неаллельное взаимодействие генов: кодоминирование, комплементарность, доминантный эпистаз, полимерию. Также этот урок включает пример решения задачи на плейотропное действие гена
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Решение генетических задач. Взаимодействие генов"
Аллельное взаимодействие генов.
Кодоминирование.
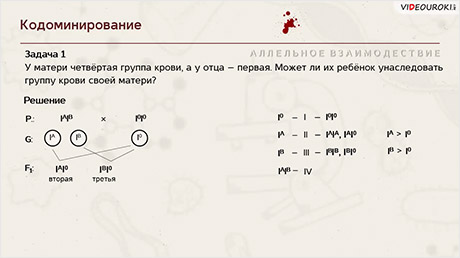
Задача 1. У матери четвёртая группа крови, а у отца – первая. Может ли их ребёнок унаследовать группу крови своей матери?
Решение: вспомним обозначения и комбинации генов, определяющие фенотипические признаки при наследовании групп крови у человека.

Возвращаемся к задаче.
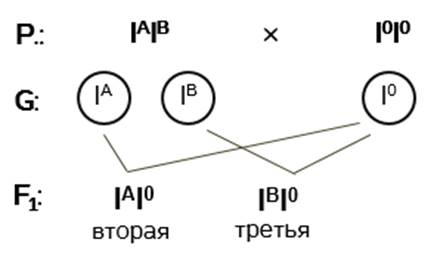
Записываем генотипы родительских форм. У матери – I A I B , у отца – I 0 I 0 .
Определяем гаметы, которые дают родительские организмы. Поскольку от гомогаметного отца ребёнок не может получить ни ген I A , ни ген I B , то у него не может быть четвёртой группы крови. У половины потомков этих родителей может быть только вторая группа крови, а у другой половины – только третья.
Ответ: нет, не может.
Задача 2. В родильном доме перепутали двух малышей. Первая пара родителей имеет I и II группы крови, вторая пара – II и IV. У одного из детей – II группа крови, а у второго – I группа. Помогите медперсоналу определить родителей обоих детей.
Решение: рассмотрим варианты генотипов первого ребёнка со второй группой крови. Это могут быть I A I A или I A I 0 . Анализируем генотипы родителей. И делаем вывод, что родителями ребёнка может быть как одна пара, так и другая.

Со вторым ребёнком всё значительно проще. Он гомогаметен, поскольку имеет первую группу крови. Значит, у обоих его родителей в генотипе должен быть ген I 0 . Такой вариант возможен только с первой парой родителей.
Ответ: ребёнок со второй группой крови – наследник второй пары родителей; ребёнок с первой группой крови рождён первой парой родителей.
Неаллельное взаимодействие генов.
Комплементарность (дополнительное взаимодействие).
Задача 3. У душистого горошка окраска цветков проявляется только при наличии двух доминантных генов: А и В. Если в генотипе имеется только один доминантный ген, то окраска не развивается. Какое потомство в первом и втором поколении получится от скрещивания растений с генотипами ААbb и ааВВ?
Решение: из условия задачи видно, что гены А и B отвечают за образование пигмента и его проявление. Но это происходит только у тех особей, в генотипе которых они встречаются вместе. В остальных случаях окраска не проявляется и цветки остаются белыми.
Записываем данные нам генотипы родительских форм. По фенотипу это два белых растения. Поскольку они гомозиготы, то образуют по одному типу гамет: Ab и aB.
В первом поколении получаем единообразие гибридов: 100% красноцветковых дигетерозигот.

Скрещиваем гибридов первого поколения между собой.
Дигетерозиготные особи дают по четыре типа гамет.
Строим решётку Пеннета.
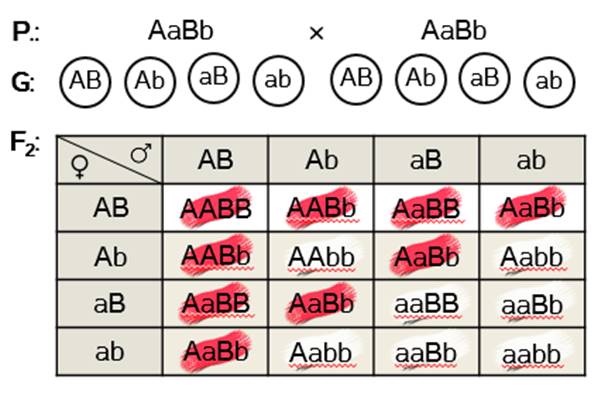
Во втором поколении гибридов получаем два варианта потомков по фенотипу – крас-ные и белые. Девять красных и семь белых. Девять к семи – это одно из соотношений расщепления, характерное для комплементарного взаимодействия генов.
Ответ: в первом поколении гибридов все особи с красными цветками дигетерозиготные. Во втором – наблюдаем расщепление: девять красноцветковых растений к семи белоцветковым.
Задача 4. При скрещивании растений одного из сортов тыквы с жёлтыми и белыми плодами всё потомство первого поколения имело белые плоды. При скрещивании гибридов F1 между собой во втором поколении было получено: 204 растения с белыми плодами, 53 – с жёлтыми и 17 – с зелёными. Определите генотип родителей и тип наследования.
Решение: определяем тип наследования. Для этого составляем соотношение расщепления.

Получаем 12:3:1. Такая пропорция говорит нам о том, что это эпистаз.
Вводим буквенные обозначения генов и записываем признаки, которые эти гены определяют. Пускай А – ген, определяющий жёлтую окраску, а – зелёную. Ген B подавляет образование пигмента, а b – не оказывает влияния на окраску.
Расщепление во втором поколении 12:3:1 означает, что родительские формы этих гибридов были дигетерозиготными, поскольку дали четыре типа гамет. А так как в первом поколении гибридов наблюдалось их единообразие, значит родительские формы гибридов первого поколения были гомозиготами по двум признакам. Одни из аллелей этих гомозигот доминантные, а другие – рецессивные.
Записываем генотипы родительских форм гибридов первого поколения: AAbb и aaBB. Проверяем соответствие записанных генотипов с фенотипами, данными по условию задачи. Одни родительские формы жёлтые, другие – белые.
Получаемое первое поколение гибридов – белые дигетерозиготы.

Скрещиваем их между собой.
Получаем заданное расщепление по фенотипу у гибридов второго поколения.
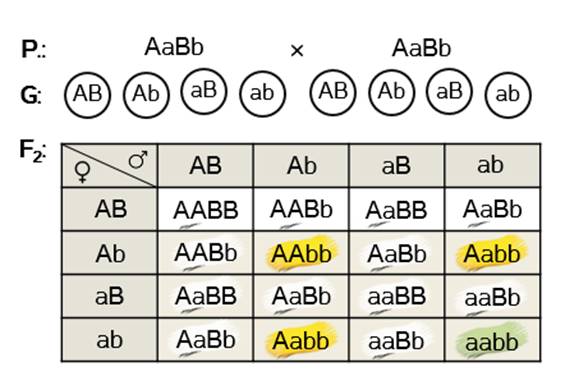
Записываем ответ задачи. Родители: доминантная гомозигота по первому признаку и рецессивная гомозигота по второму, и рецессивная гомозигота по первому признаку, доминантная гомозигота по второму. Тип наследования – доминантный эпистаз.
Задача 5. Рост человека определяется взаимодействием нескольких пар генов: А1a1A2a2А3a3. Люди с генотипом a1a1a2a2a3a3 имеют рост 150 см. С генотипом А1А1A2A2А3А3 – около 180 см (каждый доминантный ген добавляет к росту 5 см). Племя людей низкого роста порабощается ордой воинов с ростом в 180 см. Победители убивают мужчин и женятся на их женщинах. Какой рост будет у детей первого поколения от этих браков?
Решение: все гены отвечают за рост человека.
Записываем генотипы родителей. По условию – это доминантные и рецессивные гомозиготы.
Гомозиготы дают по одному типу гамет. В результате гибриды первого поколения будут гетерозиготами по трём парам генов.
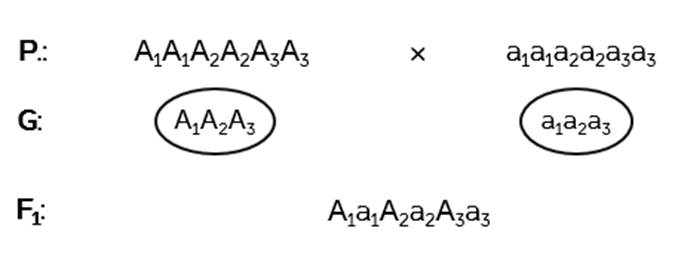
Поскольку каждый доминантный ген прибавляет к самому малому росту в 150 сантиметров по 5 см, а доминантных гена в данном случае три, значит потомки будут иметь рост 165 сантиметров.
Ответ: 165 см.
Плейотропия.
Задача 6. У мышей ген доминантной жёлтой пигментации шерсти обладает гомозиготным летальным действием. Его аллель определяет рецессивную чёрную пигментацию и обеспечивает жизнеспособность мышей. Скрещены две жёлтые особи. Какое расщепление по окраске шерсти ожидается у гибридов первого поколения?
Решение: вводим буквенные обозначения генов. Пускай ген А обуславливает жёлтую окраску шерсти, а ген а – чёрную.
При этом отмечаем, что в гомозиготном состоянии доминантные гены приводят к нежизнеспособности потомства.
Записываем генотипы родительских форм.
Жёлтые жизнеспособные мыши могут быть только гетерозиготами.
Записываем генотипы гибридов первого поколения.

Получаем четыре части гибридов, из которых одна часть доминантных гомозигот оказывается нежизнеспособной.
Ответ: живыми в потомстве окажутся две части мышей с жёлтой шерстью и одна часть – с чёрной.
Читайте также: