
После опрыскивания растений раствором абсцизовой кислоты устьица
Добавил пользователь Алексей Ф. Обновлено: 19.09.2024
Абсцизовая кислота (англ. ABA ), абсцизин, дормин — это гормон растений (изопреноид). Показана роль абсцизовой кислоты во многих процессах развития растений, в частности, в спячке почек.
Содержание
Функции
Первоначально для абсцизовой кислоты предполагалась роль в опадании листьев, но в настоящий момент такая роль показана лишь для небольшого числа растений. У растений описаны сигнальные пути связанные с абсцизовой кислотой при ответе на стресс и патогены. [2] [3] Обнаружены и секвенированы гены, продукты которых принимают участие в биосинтезе абсцизовой кислоты. [4] [5] Абсцизовая кислота синтезируется некоторыми патогенными грибами по другому пути, чем у растений. [6]
Показана роль абсцизовой кислоты в опадании листьев. При подготовке к зиме абсцизовая кислота синтезируется в концевых почках растений. Это приводит к замедлению роста, а из прилистников образуются защитные чешуйки, покрывающие спящие почки в холодный период. Абсцизовая кислота останавливает деление клеток камбия и останавливает первичный и вторичный рост.
Абсцизовая кислота также образуется в корнях растений в ответ на снижение водного потенциала, а также при стрессе. Затем абсцизовая кислота поступает в листья, где изменяет осмотический потенциал устьичных клеток, и вызывает закрывание устьиц. Закрывание устьиц снижает транспирацию и предотвращает дальнейшую потерю воды через листья.
Описаны мутанты Arabidopsis thaliana, дефицитные по синтезу абсцизовой кислоты. Такие растения имеют нарушения покоя семян, при прорастании, в структуре устьиц, некоторые мутанты имеют задержки роста и коричнево-желтые листья. [7]

Гормональная система растений Сигналы о стрессе

Абсцизовая кислота (АБК) англ. dormancy – покой лат. abscission – опадение Дормин Абсцизин

Абсцизовая кислота (АБК) Биосинтез абсцизовой кислоты Мутанты кукурузы из серии viviparous: vp2 – vp10

Абсцизовая кислота (АБК) 1.Зеаксантин 2. Антераксантин 3. Виолаксантин 4. all-trans-неоксантин 5. цис-неоксантин 6. Ксантоксин 7. Абсцизовый альдегид 8. Абсцизовая кислота (АБК) Биосинтез абсцизовой кислоты
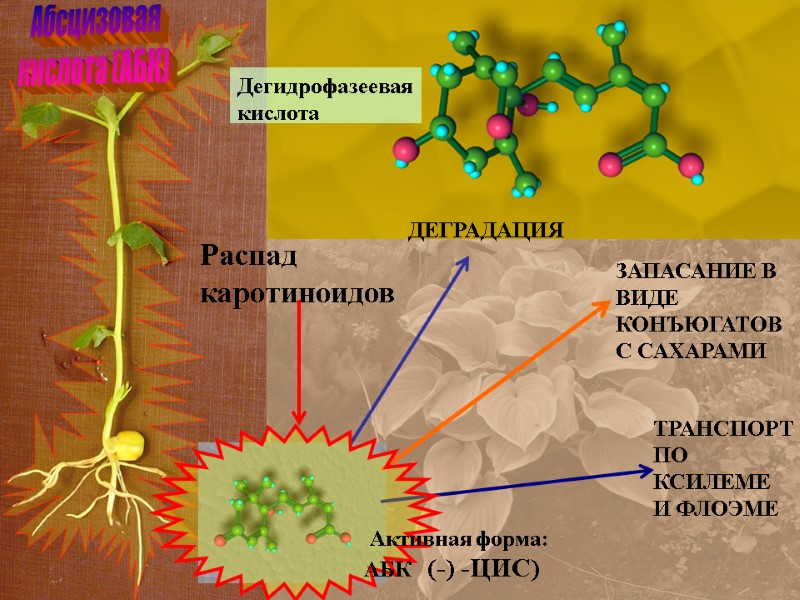
Абсцизовая кислота (АБК) Распад каротиноидов ДЕГРАДАЦИЯ ТРАНСПОРТ ПО КСИЛЕМЕ И ФЛОЭМЕ ЗАПАСАНИЕ В ВИДЕ КОНЪЮГАТОВ С САХАРАМИ Дегидрофазеевая кислота Активная форма: АБК (-) -ЦИС)
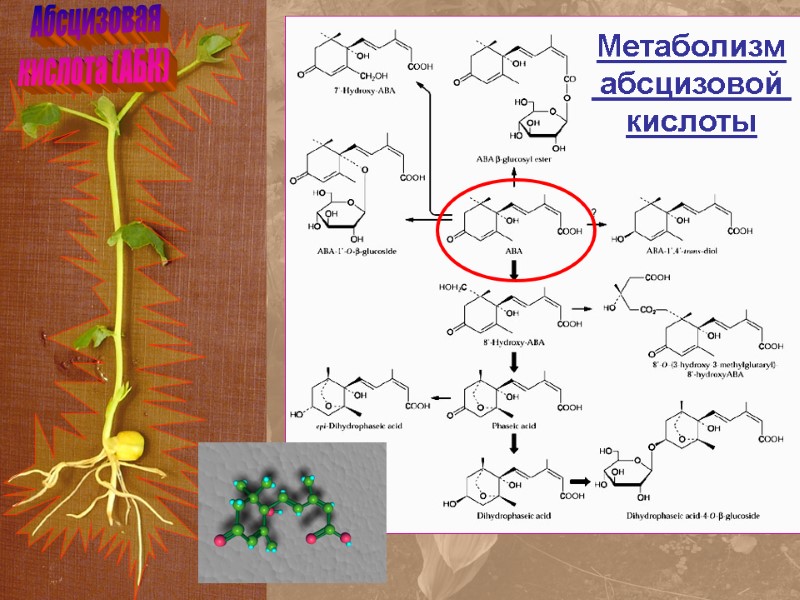
Абсцизовая кислота (АБК) Метаболизм абсцизовой кислоты

Абсцизовая кислота (АБК) Сигнальная система АБК, регулирующая время цветения – внутриклеточный рецептор. FCA (Flowering time Control A) – РНК-связывающий ядерный белок, который запускает цветение, снимая действие FLC (Flowering Locus C) - мощного ингибитора цветения. FCA афинно и специфично связывется с АБК. Без АБК, FCA образует совместно с фактором полиаденилирования РНК FY (Flowering Locus Y) комплекс процессинга РНК, который предотвращает образование mРНК ингибитора FLC. Растение цветет. При наличии АБК она связывается c С-концевым участком FCA, рядом с местом взаимодействия с FY. Это разрушает их взаимодействие, что ведет к накоплению FLC и задержке цветения. Транскрипция SOC1 происходит только в растениях с низкими уровнями FLC

Абсцизовая кислота (АБК) По коллекции мутантов Arabidopsis, не чувствительных к АБК (ABA-insensitive – ABI ) найдены компоненты системы трансдукции сигнала: ABI1: серин-треониновая протеин-фосфатаза, 434а-к, имеет Са2+-связывающий участок. ABI2 – 80% гомологии с ABI1. ABI3 – образует комплексы с факторами транскрипции. ABI5 – связывается с ABRC-мотивом в ДНК промоторов. Гены-мишени: сдержат ABRE или ABRC (АБК-регулируемый элемент или комплекс). В промоторной области - G-бокс, обычно с последовательностью ACGT Регуляторные механизмы и трансфакторы АБК-сигналинга у Arabidopsis thaliana = vp1 (Zea)
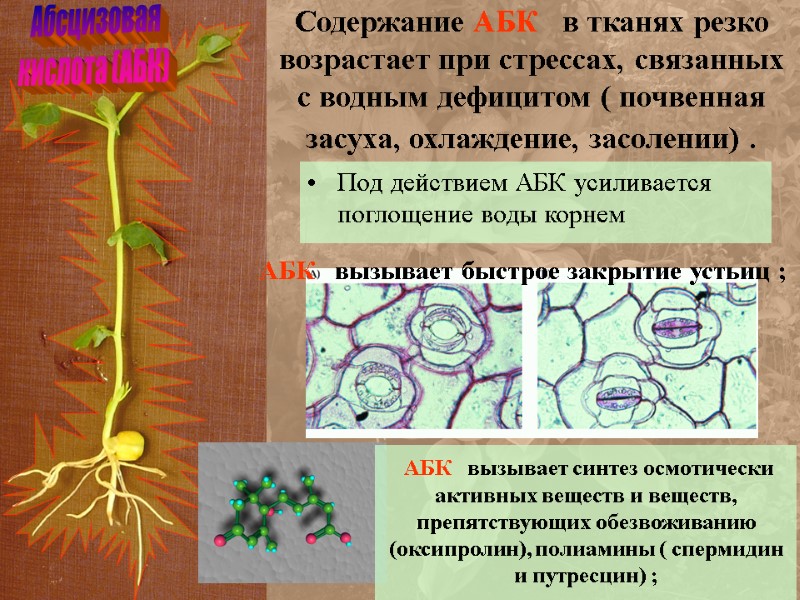
Абсцизовая кислота (АБК) Содержание АБК в тканях резко возрастает при стрессах, связанных с водным дефицитом ( почвенная засуха, охлаждение, засолении) . АБК вызывает быстрое закрытие устьиц ; Под действием АБК усиливается поглощение воды корнем АБК вызывает синтез осмотически активных веществ и веществ, препятствующих обезвоживанию (оксипролин), полиамины ( спермидин и путресцин) ;

Реакция устьиц на абсцизовую кислоту и цитокинины Цитокинины Абсцизовая кислота Цитокинин Абсцизовая кислота (АБК)
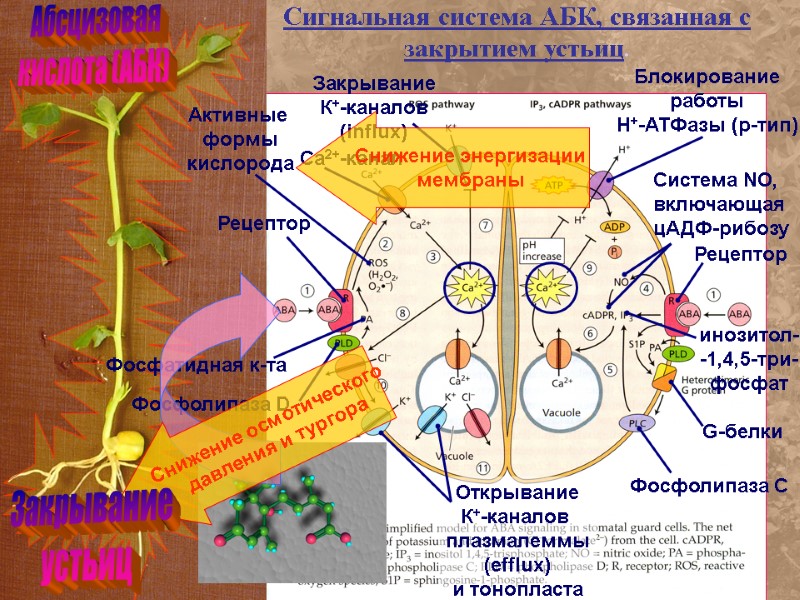
Абсцизовая кислота (АБК) Сигнальная система АБК, связанная с закрытием устьиц. Рецептор Фосфолипаза D Фосфатидная к-та Активные формы кислорода Са2+-канал Открывание К+-каналов плазмалеммы (efflux) и тонопласта Закрывание К+-каналов (influx) Рецептор G-белки Фосфолипаза С Система NO, включающая цАДФ-рибозу инозитол- -1,4,5-три- фосфат Блокирование работы Н+-АТФазы (р-тип) Снижение осмотического давления и тургора Закрывание устьиц Снижение энергизации мембраны

Абсцизовая кислота (АБК) Адаптация к низким температурам Повышается устойчивость к низким температурам

Абсцизовая кислота (АБК) Переход в состояние покоя Остановка роста 2. Запасание питательных веществ 3. Синтез осмопротекторных соединений (они же – антифризы) 4. Образование пробковой ткани (перидерма) 5. Сброс листьев 7. Дифференциальный переход ДНК в более метилированное и конденсированное состояние 6. Образование специализированных органов (катафиллы, клубни, выводковые почки и др.)

Абсцизовая кислота (АБК) Окончание физиологического покоя

Абсцизовая кислота (АБК) Физиологический покой семян Стратификация холодовая обработка набухших семян при околонулевых температурах 2. Вымывание веществ-ингибиторов талыми водами
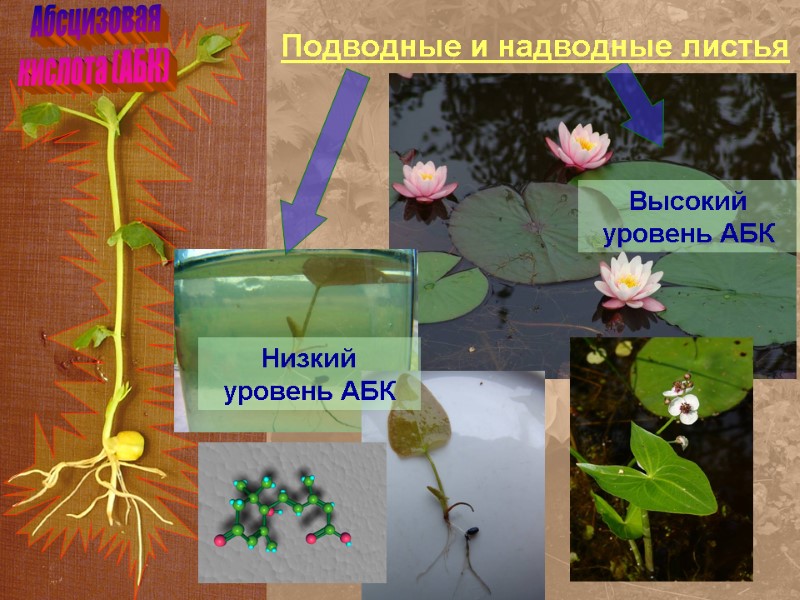
Абсцизовая кислота (АБК) Подводные и надводные листья Высокий уровень АБК Низкий уровень АБК

Абсцизовая кислота (АБК) Патогенные грибы синтезируют АБК Botrytis cinerea

Механический стресс Этилен Апикальная петелька Утолщение
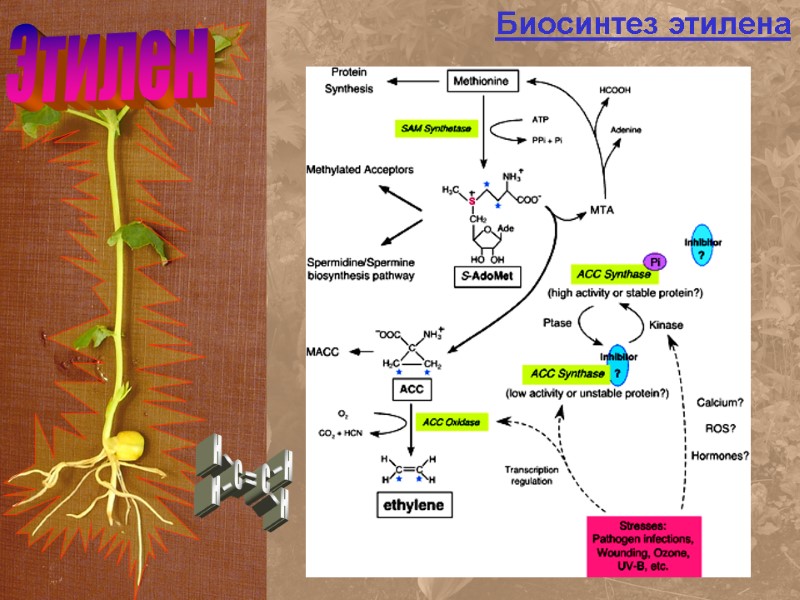
Этилен Биосинтез этилена

Этилен Биосинтез этилена Предшестенник этилена (АЦК) Образуется в корнях и транспортиру-ется в стебли и листья В аэробной среде из АЦК образуется этилен, растения вянут

Этилен Мутанты, не чувствительные к этилену

Этилен Этилен и корневые волоски

Этилен Рецепция этилена

Этилен Рецепция этилена

Этилен Рецепция этилена

Этилен Нападение вредителей

Этилен Нападение грибов

Этилен Созревание сочных плодов 1. Активация пектиназ в апоплоасте (размягчение ткани плода) 2. Распад кислот, терпких веществ в вакуоли, распад крахмала 3. Накопление в вакуоли растворимых сахаров 4. Распад хлорофилла 5. Синтез окрашенных соединений (каротиноиды и/или антоцианы) 6. Действие на зону опадения

Этилен Созревание сочных плодов + Нападение вредителей Ускоренное созревание Действие на зону опадения

Этилен Созревание сочных плодов + Нападение грибов

Этилен Этилен и опыление Увядание и опадение привлекающего аппарата (лепестки, листочки околоцветника и др.) Увядание тычинок Активный рост завязей

Этилен Цветение ананасов

Этилен Другие бромелиевые

Этилен Ароидные

О СООН ЖАСМОНОВАЯ КИСЛОТА Образуется в результате гидролиза мембранного фосфолипида фосфолипазой А. Образует летучее производное метилжасмонат (10-6- 10 -7 М)

Структура и метаболизм жасмонатов
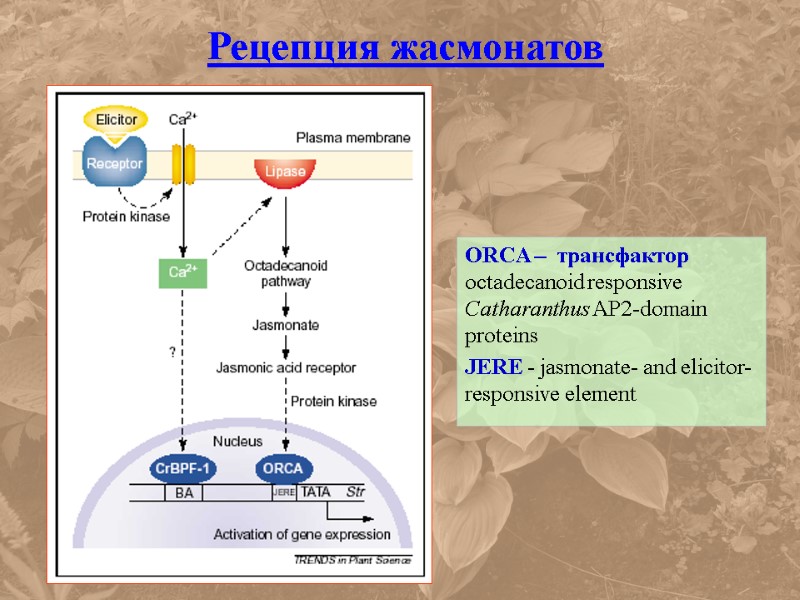
ORCA – трансфактор octadecanoid responsive Catharanthus AP2-domain proteins JERE - jasmonate- and elicitor-responsive element Рецепция жасмонатов

Эффекты жасмоновой кислоты Ингибирует рост растяжением у проростков и отрезков стеблей Ингибирует прорастание пыльцевых трубок Ингибирует рост корней Подавляет прорастание семян Вызывает в зародышах синтез запасных белков семени и белков позднего эмбриогенеза

Вторичный метаболизм (Фенольные соединения) Лирическое отступление

Фенольные соединения классифицируют по количеству ароматических колец и числу дополнительных углеродных атомов. С6 – С1 соединения (фенолокислоты) С6 – С3 соединения (оксикоричные кислоты, кумарины)
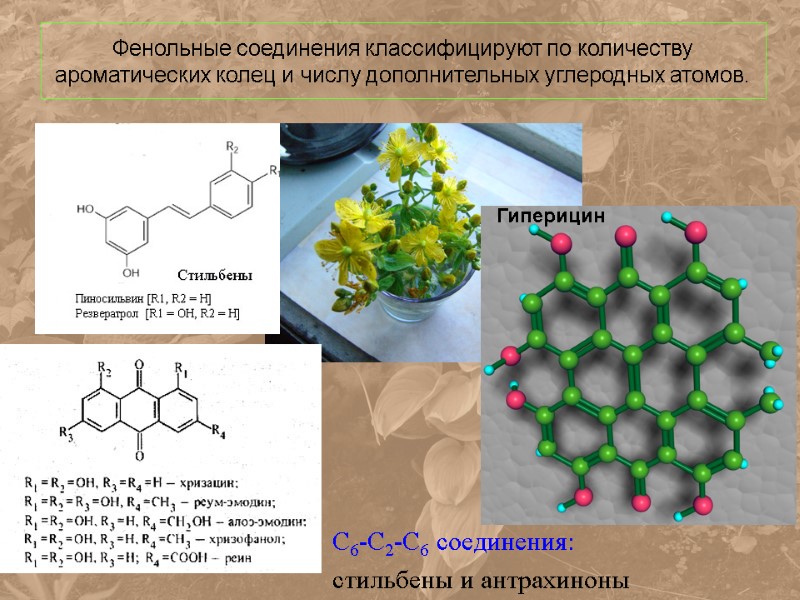
Фенольные соединения классифицируют по количеству ароматических колец и числу дополнительных углеродных атомов. С6-С2-С6 соединения: стильбены и антрахиноны
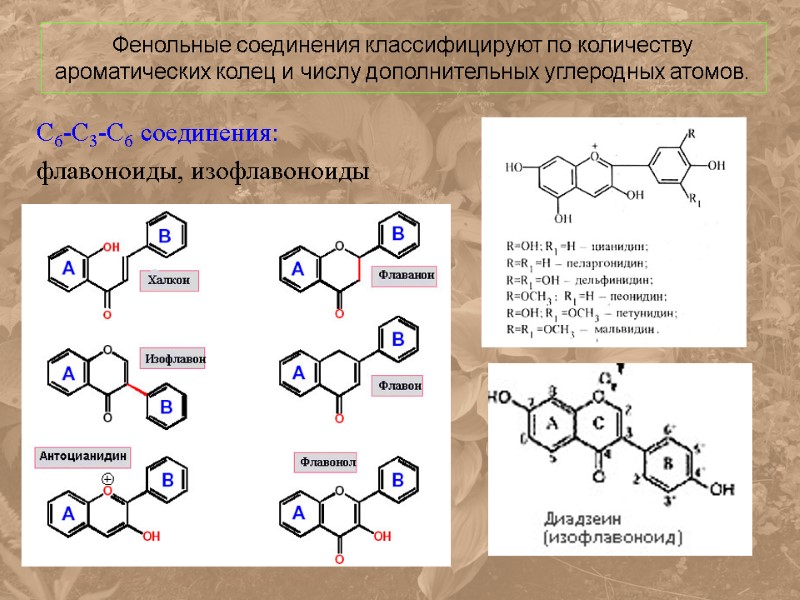
Фенольные соединения классифицируют по количеству ароматических колец и числу дополнительных углеродных атомов. С6-С3-С6 соединения: флавоноиды, изофлавоноиды

Вся радуга окраски цветков обусловлена гликозидами трех агликонов – но с модификациями

Красная и синяя окраска - АНТОЦИАНЫ Heucherella ‘Color Glory’ Heuchera ‘Snow Scrolls’ Кислая среда Щелочная среда Arum maculatum

Петунидин Пеонидин Цианидин Пеларгонидин Дельфинидин

Антоцианы – гликозиды антоцианидинов Цвет антоцианов определяется многими факторами…
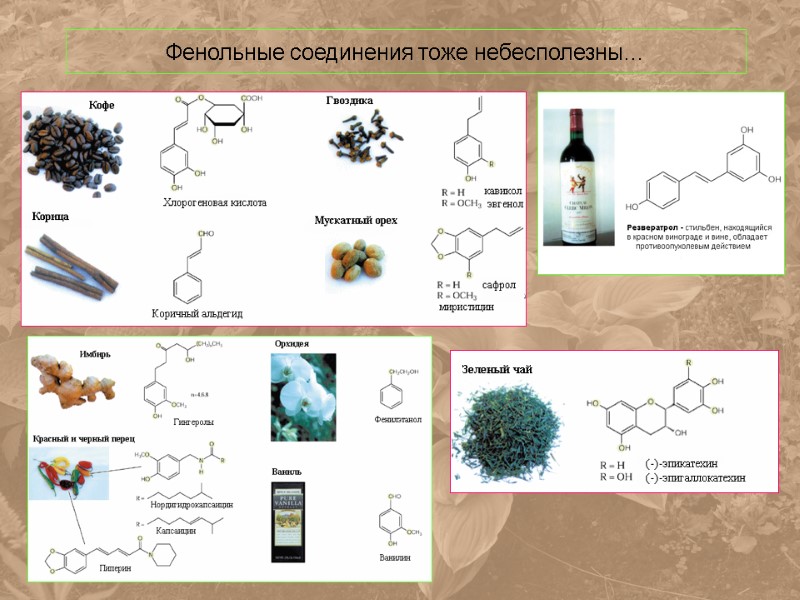
Фенольные соединения тоже небесполезны…
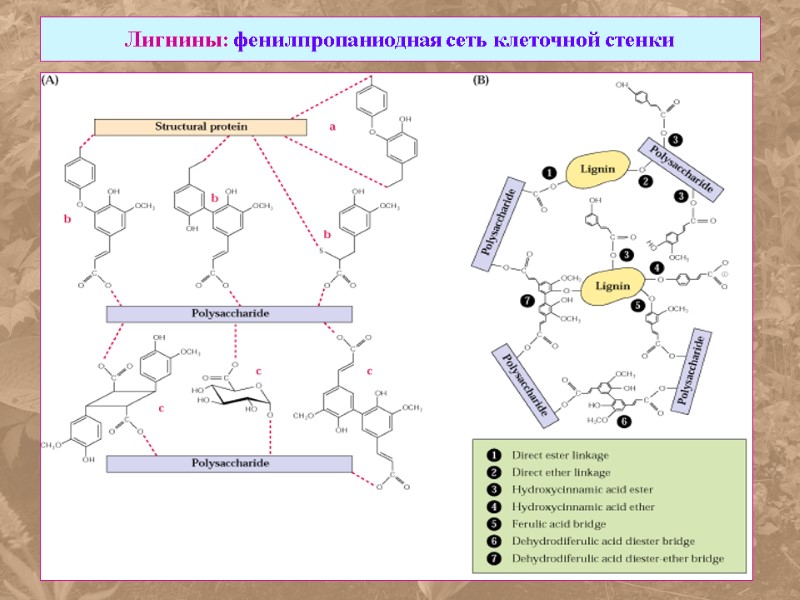
Лигнины: фенилпропаниодная сеть клеточной стенки

Образование лигнина: окислительная конденсация фенилпропаноидов случайным образом.

Образование лигнинов: целенаправленная конденсация мономеров.

Фитопатология

Этилен, салициловая кислота, олигосахарины, жасмоновая кислота участвуют в ответе растения на биотический стресс и возникновении системной устойчивости Проникновение гриба Rhynchosporium secalis после прорастания спор через поверхность листа ячменя

Атака патогена и защитные реакции
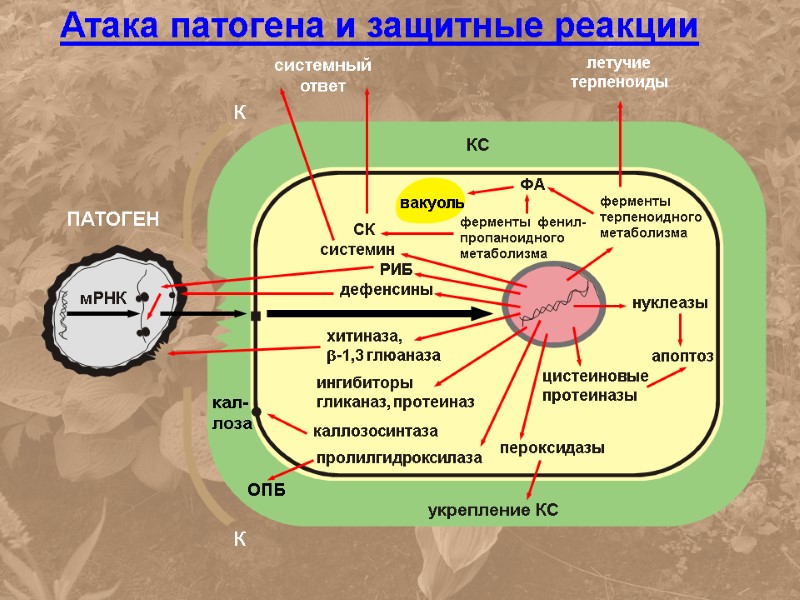
ПАТОГЕН системин ферменты терпеноидного метаболизма ФА СК ингибиторы гликаназ, протеиназ пролилгидроксилаза каллозосинтаза летучие терпеноиды цистеиновые протеиназы нуклеазы апоптоз РИБ дефенсины хитиназа, b-1,3 глюаназа пероксидазы вакуоль системный ответ ферменты фенил-пропаноидного метаболизма мРНК К К укрепление КС ОПБ кал- лоза КС Атака патогена и защитные реакции
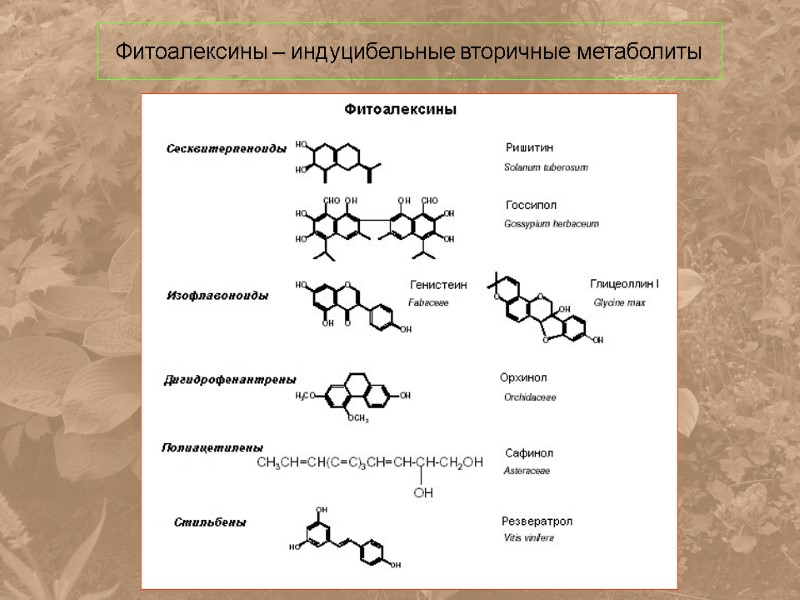
Фитоалексины – индуцибельные вторичные метаболиты
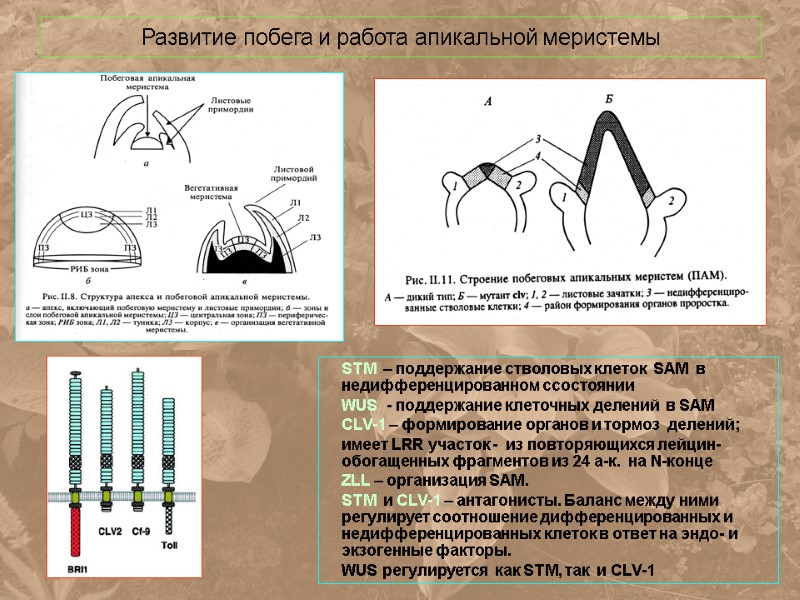
Развитие побега и работа апикальной меристемы STM – поддержание стволовых клеток SAM в недифференцированном cсостоянии WUS - поддержание клеточных делений в SAM CLV-1 – формирование органов и тормоз делений; имеет LRR участок - из повторяющихся лейцин-обогащенных фрагментов из 24 а-к. на N-конце ZLL – организация SAM. STM и CLV-1 – антагонисты. Баланс между ними регулирует соотношение дифференцированных и недифференцированных клеток в ответ на эндо- и экзогенные факторы. WUS регулируется как STM, так и CLV-1

Развитие SAM регулируется по типу отрицательной обратной связи WUSCHEL (WUS) – кодирует трансфактор с гомеодоменом. Активизирует деление клеток SAM CLAVATA (CLV) 1,2,3, - функционально связанные гены. CLV1 – LRR киназа, CLV2 – LRR-белок. CLV3 – небольшой (11 kDa) водорастворимый белок. KAPP – фосфатаза.
Абсци́зовая кислота́ (англ. ABA ), абсцизин, дормин — это гормон растений (изопреноид) [2] . Впервые была обнаружена в экспериментах по поиску вещества по способности вызывать опадение листьев и коробочек хлопчатника. Первые препараты абсцизовой кислоты (АБК) были независимо выделены в 1963 г. из листьев берёзы Ф. Эддикоттом и сотрудниками (США) и Ф. Уорингом и сотрудниками (Великобритания) [3] .
Основное место синтеза АБК — листья и корневой чехлик. Она присутствует в клетке как в свободной форме, так и в виде конъюгатов с глюкозой [4] .
АБК называют гормоном-антагонистом ауксина, цитокининов, гиббереллина, так как она тормозит реакции, которые вызывают эти гормоны [4] .

Лекции
Лабораторные
Справочники
Эссе
Вопросы
Стандарты
Программы
Дипломные
Курсовые
Помогалки
Графические
Доступные файлы (6):
(6) Ингибиторы роста -АБК и этилен.doc
^ Физиологические проявления действия абсцизовой кислоты АБК тормозит процессы роста, индуцированные ИУК, цитокинином и гиббереллином. Накопление АБК приводит к снижению фотосинтетического фосфорилирования и интенсивности фотосинтеза. Увеличение содержания АБК тормозит рост пазушных почек при апикальном доминировании, задерживает прорастание семян, влияет на переход в покоящееся состояние семян, почек, клубней. Обычно она накапливается перед наступлением зимних холодов, а ко времени окончания покоя ее содержание уменьшается. Ингибиторное действие АБК на прорастание семян и рост тканей в ряде случаев может сниматься обработкой гиббереллинами или цитокининами. АБК регулирует опадение листьев и плодов. Обработка АБК вызывает старение и опадение листьев у ряда растений. АБК способствует образованию запасных белков, выступает антагонистом в индукции гиббереллином синтеза а-амилазы, а также вызванного цитокинином роста активности нитратредуктазы. Действие АБК показано на уровне регуляции экспрессии генов. Подавление ею роста связано с репрограммированием генома и синтезом большого числа АБК-индуцируемых полипептидов. АБК рассматривают как антистрессорный фактор, усиливающий адаптацию растений к различным неблагоприятным воздействиям. В условиях водного стресса наблюдается быстрое увеличение содержания АБК. Перераспределение АБК: она транспортируется из клеток мезофилла в замыкающие клетки устьиц. При этом возрастает проницаемость мембран, что приводит к утечке ионов К+ из замыкающих клеток, осмотическая концентрация снижается и вода выходит, устьица закрываются. Реакция закрывания устьиц на АБК является одной из самых быстрых известных реакций на действие фитогормона, так как обычно она происходит в течение 5—10 мин. Одновременно с закрытием устьиц снижается интенсивность транспирации. Полагают, что задержка ростовых процессов при засухе также связана с накоплением АБК в клетках апекса, куда она перетекает из листьев. С уменьшением расходования воды АБК при засухе усиливает поглощение воды корневой системой, стимулирует пасокодвижение. Таким образом, обработанные АБК растения в условиях водного дефицита характеризуются более высоким содержанием воды. Обработка АБК способствует повышению устойчивости к засухе, затоплению, высоким и низким температурам, морозу, солям. Это связано с действием АБК на водный баланс растений, фотосинтез и дыхание, обмен липидов, стабилизацией цитоскелетных структур, модификацией проницаемости клеточных мембран, торможением роста. АБК ускоряет синтез протекторных белков, определяющих устойчивость к неблагоприятным условиям. На поздних стадиях эмбриогенеза повышается содержания АБК и появляются специфические РНК и соответствующие белки. Это так называемые белки позднего эмбриогенеза, которые участвуют в обеспечении устойчивости к обезвоживанию. Обработка АБК повышает содержание этих белков. Большую роль в действии АБК играют процессы фосфорилирования и дефосфорилирования, что связано с работой протеинкиназ. В опытах с инъекциями показано, что действие АБК на движение устьиц не проявляется при нарушении процессов фосфорилирования и дефосфорилирования, а также при торможении работы протеинкиназ.
^ Этилен — это газ. Химическая формула СН2= СН2. Этилен отнесен к фитогормонам сравнительно недавно. Однако еще в 1911 г. русский ученый Д.Н. Нелюбов установил, что этилен тормозит рост стебля в длину, одновременно вызывая его утолщение и изгиб в горизонтальном направлении (тройная реакция стебля). В последующем было показано, что сочные плоды ряда растений (апельсины, бананы и др.) выделяют этилен, и что он стимулирует созревание плодов. В 1935— 1937 гг. Хичкок и Циммерман в США и Ю.В. Ракитин в СССР провели большое количество исследований, показавших, что этилен — регулятор созревания плодов. В 60-е годы показано, что спектр действия этилена значительно шире и что, подобно АБК, этот фитогормон оказывает в основном тормозящее влияние на процессы роста. Первоначальным предшественником образования этилена является аминокислота метионин. В образовании этилена участвует ряд ферментов, из которых особое значение имеет аминоциклопропанкарбосинтаза (АЦК-синтаза), катализирующая образование 1-аминоциклопропан-1-карбоновой кислоты (АЦК) — непосредственного предшественника этилена. Активность этого фермента возрастает в процессе созревания плодов, при поранении и, что самое главное, регулируется ауксином. Показано, что высокие концентрации ауксина вызывают синтез этилена. Предполагают, что происходит на уровне индукции генов АЦК-синтазы. Этилен образуется в созревающих плодах, стареющих листьях, в проростках до того, как они выходят на поверхность почвы. В растении этилен определяют с помощью биотестов или газовой хроматографии.
^ Физиологические проявления действия этилена
Наиболее яркое проявление действия этилена — это регуляция процессов созревания плодов. Плоды в период созревания образуют этилен, который и регулирует все эти процессы. Ингибиторы синтеза этилена задерживают созревание плодов. Этилен способствует увеличению толщины, но уменьшает рост в длину стебля, а также клеток, что связано с изменением ориентации микрофибрилл целлюлозы. Способствует образованию отделительного слоя и опадению листьев и плодов. Образование отделительного слоя связано с появлением ферментов, растворяющих клеточные стенки, нарушением связей между клетками. Этилен ускоряет процессы старения, тормозит рост почек, накапливается в покоящихся органах. Во многих случаях его накопление и действие связано с ауксином. Ауксин в повышенной концентрации вызывает образование этилена и, как следствие, торможение ростовых процессов. Возможно, торможение роста, вызванное высокими концентрациями ауксина, связано с накоплением этилена. Так, показано, что этилен и высокие концентрации ауксина вызывают эпинастию листьев, т. е. изменение угла наклона листа по отношению к стеблю в результате чего листья опускаются. У некоторых растений (ананасы) этилен индуцирует образование цветков. Этилен влияет на пол цветков, вызывая образование женских цветков у однодомных растений (огурец, тыква). При затоплении растений этилен индуцирует образование корней на стебле и формирование аэренхимы — ткани стебля, по которой кислород поступает в корни. Это позволяет растениям выживать в условиях кислородного голодания корней. Кроме того, этилен индуцирует образование на стебле адвентивных корней. Эти корни не выполняют поглощающую функцию, а участвуют в снабжении побегов веществами, необходимыми для нормального функционирования, например цитокининами. Этилен участвует в реакции растений на повреждающие воздействия, в частности на патогенные микроорганизмы (грибы, бактерии, вирусы). Под действием этилена в растении синтезируются белки-ферменты, такие как хитиназа и глюканаза, которые разрушают клеточную стенку патогенов. Есть данные, что этилен индуцирует и синтез ферментов, участвующих в образовании защитных соединений, например фитоалексинов.
^ Взаимодействие фитогормонов Рост растений и формообразовательные процессы регулируются определенным соотношением фитогормонов. Значение соотношений между ауксином и цитокинином хорошо показано в опытах с выращиванием изолированных тканей. Увеличение в питательной среде отношения ауксин/кинетин приводит к тому, что из массы недифференцированных клеток (каллуса), выращиваемых в стерильных условиях, дифференцируется корень. Уменьшение указанного соотношения приводит к дифференциации побегов. Нельзя также не учитывать, что одни гормоны могут оказывать влияние на ферменты, катализирующие синтез или разрушение других гормонов, и тем самым изменять их содержание. Так, известно, что под влиянием гиббереллинов содержание ауксинов растет, ауксин может вызывать накопление этилена. Каждый из фитогормонов играет свою специфическую роль в процессах обмена, внешнее проявление их действия может быть одинаковым (рост клеток растяжением, деление клеток).
^ Применение фитогормонов в практике растениеводства
Для практического применения фитогормонов или их синтетических заменителей необходимо соблюдение определенных условий:
1. Фитогормоны оказывают влияние лишь тогда, когда в растении их недостает. Это чаще всего наблюдается во время прорастания семян, цветения, образования плодов, а также когда нарушена целостность растительного организма, например, черенки, изолированные ткани. В некоторых случаях условия внешней среды препятствуют образованию гормонов, тогда этот недостаток может быть восполнен их экзогенным внесением. 2. Как уже упоминалось, клетки, ткани, органы должны быть компетентны (восприимчивы) к фитогормонам. Компетенция связана с наличием рецепторов, а также с общим состоянием внутриклеточных процессов. Клетка может быть на одной фазе роста компетентна к внесению данного фитогормона, а на другой — нет. 3. Необходимо достаточное снабжение растения водой и питательными веществами. 4. Действие всех гормонов зависит от концентрации. Избыточная концентрация вызывает не стимуляцию, а резкое торможение роста и даже гибель растений.
5.Эндогенные (естественные) фитогормоны определенным образом локализованы в отдельных компартментах (отсеках) клетки. При внесении извне распределение гормонов будет иным. В этой связи экзогенное внесение не может полностью заменить гормоны, образовавшиеся при естественном метаболизме.
В настоящее время фитогормоны применяются в следующих направлениях.
^ Ауксины и их синтетические заменители
1. Для усиления корнеобразования у черенков. Обработка нижних концов черенков ИУК в концентрации 50 мг/л или ее синтетическими аналогами (индолилмасляной, а-нафтилуксусной кислотой) вызывает приток к ним питательных веществ, при этом процесс корнеобразования усиливается. Этот прием широко применяется при вегетативном размножении растений. 2. Для усиления корнеобразования и восстановления корневой системы при пересадке растений. Для этого саженцы окунают в пасту из глины с добавлением ИУК или ИМК. 3. Для образования партенокарпических плодов, повышения урожая томатов и некоторых других культур. Опрыскивание цветков томатов раствором синтетических регуляторов роста типа ауксина (например, трихлорфеноксиуксусной кислотой в концентрации 50 мг/л) приводит к образованию партенокарпических бессемянных плодов. Плоды растут быстрее и характеризуются более высоким содержанием Сахаров. Одновременно с усилением роста плодов в результате перераспределения питательных веществ рост вегетативных органов (пасынков) замедляется. К недостаткам этого приема следует отнести большую подверженность образующихся плодов различного рода заболеваниям. 4. Для предохранения плодов от предуборочного опадения. При большом количестве завязавшихся плодов часть их опадает до созревания. Регуляторы типа ауксина, вызывая дополнительный приток питательных веществ к плодам, препятствуют образованию отделительного слоя. Обработка деревьев проводится а-нафтилуксусной кислотой в концентрации 10 мг/л за две недели до уборки. 5. Для ускорения прорастания семян некоторых растений. Этот прием дает благоприятные результаты лишь на мелкосемянных растениях, поскольку крупные семена содержат достаточное количество собственных гормонов. Хорошие результаты получены при обработке семян сахарной свеклы ИУК в концентрации 10 мг/л. 6. В высоких концентрациях регуляторы роста типа ауксина, например 2,4-дихлорфеноксиуксусная кислота (2,4-Д), могут применяться как селективные гербициды. Поскольку для разных видов растений оптимальные концентрации фитогормонов различны, то это позволяет использовать 2,4-Д в качестве селективного гербицида для борьбы с сорняками в посевах злаковых культур. Показано, что различная устойчивость растений к 2,4-Д связана с различиями в скорости ее детоксикации (обезвреживания) в растительном организме.
Гиббереллины.
1. Под влиянием гиббереллина заметно усиливается рост стебля конопли, сахарного тростника. Увеличивается выход волокна конопли с гектара. 2. Опрыскивание в концентрации 25 мг/л повышает урожай зеленой массы кормовых бобов. Увеличивается продуктивность растений, возделываемых для получения зеленой массы. Однако урожай семян при этом снижается.
Широкое применение получили этиленпродуценты — соединения, которые после опрыскивания растении распадаются с образованием этилена. Наиболее распространенным этиленпродуцентом является 2-хлорэтилфосфоновая кислота СlСН2СН2РО(ОН)2, которая при значении рН > 4 распадается с выделением этилена. Известно много препаратов, содержащих 2-ХЭФК: этрел, гидрел, кампозан, этефон. Этилен-продуценты применяют:
1. Для ускорения созревания плодов. 2. Для ускорения созревания и облегчения уборки томатов, яблок, облепихи, винограда, цитрусовых, поскольку ускоряется образование отделительного слоя в плодоножке. 3. Для усиления отделения латекса у каучуконосных деревьев гевеи, ускорять выделение живицы у сосны. 4. Для ускорения опадения листьев — дефолиации. Дефолиация облегчает условия машинной уборки ряда культур, например хлопчатника. 5. Для индукции цветения манго, авокадо, ананасов и других культур. 6. Обработка этрелом растений огурца вызывает преимущественное образование женских цветков. 7. Для борьбы с полеганием зерновых культур в посевах, например, ржи и ячменя, поскольку этиленпродуценты способствуют формированию укороченного толстого стебля.


Рис. 1. Arabidopsis thaliana обычный (WT, wild type) и модифицированный, переведенный в режим экономии воды (PYR1 MANDI ), после небольшой самодельной засухи. Изображение из обсуждаемой статьи в Nature
При недостатке воды растения выделяют фитогормон — абсцизовую кислоту (АБК), — который изменяет активность генов, переводя растение в режим экономии. Однако этот гормон дорог в получении и быстро разлагается. Ученые создали вариант рецептора, который вместо АБК активируется уже используемым в сельском хозяйстве химикатом, безвредным для человека. У растений с таким рецептором режим устойчивости к засухе можно включать при первой необходимости.
Растения чутко реагируют на недостаток воды: у них выделяется фитогормон под названием абсцизовая кислота (АБК), который изменяет активность генов, переводя растение в режим экономии воды. При этом у растения уменьшается просвет устьиц в листьях, благодаря чему оно теряет меньше воды за счет испарения.
Для сельского хозяйства было бы очень удобно поставить этот процесс под контроль, чтобы у ценных растений режим экономии воды можно было включать заблаговременно, еще до того, как растение начнет чувствовать влияние засухи. Самый простой, на первый взгляд, способ помочь растениям бороться с засухой — это опрыскивать их раствором абсцизовой кислоты. Однако синтез этого вещества обходится дорого, и, кроме того, оно довольно быстро разлагается.
Изменение просвета устьиц у растений детектируют интересным косвенным способом — по температуре листьев. Испарение воды через устьица снижает температуру листьев, а если устьица прикрываются, то испарение воды уменьшается, а температура листьев растет. Ученые убедились, что у полученных модифицированных растений при обработке мандипропамидом температура листьев повышается (рис. 2). Значит, мандипропамид уменьшал просвет устьиц на листьях — то есть заставлял растения экономить воду.

Источники:
1) Masanori Okamoto, Francis C. Peterson, Andrew Defries, Sang-Youl Park, Akira Endo, Eiji Nambara, Brian F. Volkman, and Sean R. Cutler. Activation of dimeric ABA receptors elicits guard cell closure, ABA-regulated gene expression, and drought tolerance // Proceedings of the National Academy of Sciences of the United States of America. 2013. V. 110. P. 12132–12137.
2) Sang-Youl Park, Francis C. Peterson, Assaf Mosquna, Jin Yao, Brian F. Volkman & Sean R. Cutler. Agrochemical control of plant water use using engineered abscisic acid receptors // Nature. Published online 04 February 2015. Doi:10.1038/nature14123.
Читайте также: