
Образование пыльцы у покрытосеменных или папоротниковидных
Добавил пользователь Евгений Кузнецов Обновлено: 15.09.2024
ГЛАВА 8. ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ. ОБРАЗОВАНИЕ ЗАРОДЫША И ЭНДОСПЕРМА
8.1. Опыление покрытосеменных растений
Несмотря на то что образование женского гаметофита начинается значительно позже мужского, созревание гаметофитов происходит почти одновременно. К моменту созревания мужского гаметофита женский гаметофит готов к процессу оплодотворения.
У большинства покрытосеменных растений процессу оплодотворения предшествует опыление — перенос пыльцы из тычинок на рыльце пестика. В ходе эволюции сформировалось два типа опыления: автогамия (самоопыление) и аллогамия, или ксеногамия (перекрестное опыление). При самоопылении пыльца переносится с тычинок на пестик в пределах одного и того же цветка. При перекрестном опылении пыльца одного цветка переносится на рыльце пестика другого цветка. Самоопыление может наблюдаться как в открытых (хазмогамных) цветках (горох — Pisum), так и в закрытых (клейстогамных) (фиалка — Viola, кислица — Oxalis) (рис. 188). У некоторых фиалок и кислицы обыкновенной рано весной развиваются хазмогамные цветки, однако так как весной опылителей мало, эти цветки, как правило, не опыляются и остаются бесплодными. Позже у таких растений наряду с хазмогамными цветками у основания побегов развиваются невзрачные клейстогамные цветки, в которых происходит опыление и оплодотворение, и, как следствие этого, образуются плоды и семена.
Рис. 188. Хазмогамные (1) и клейстогамные (2) цветки у фиалки жестковолосистой (Viola hirta)

Особым типом опыления является гейтоногамия — перенос пыльцы с одного цветка на другой, но находящийся на одном и том же растении. Одни ученые рассматривают этот тип опыления как расширенное самоопыление, а другие — как ограниченное перекрестное опыление.
Перекрестное опыление может быть абиотическим и биотическим. Преобладающим типом абиотического опыления является ветроопыление. У растений выработался ряд приспособлений к ветроопылению (анемофилии): цветение древесных растений до распускания листьев (ольха, орешник); длинные раскачивающиеся под действием ветра соцветия-сережки (ольха, орешник) или длинные тычинки с крупными пыльниками (злаки); невзрачные раздельнополые цветки без околоцветника (осоки) или с сильно редуцированным околоцветником (ситники); большое количество легкой пыльцы с гладкой экзиной (орешник, береза) и т. п.
Древним типом абиотического опыления является гидрофилия — опыление при помощи воды. Типичная гидрофилия характерна для водных (водяная звездочка — Callitriche, наяда — Najas, роголистник — Ceratophyllum) растений, у которых существуют специфические особенности пыльцы. У некоторых гидрофильных растений пыльца сильно вытянута (у Zosteraдлина ее 2500 мкм), что имеет важное приспособительное значение, так как такая пыльца очень подвижна и способна быстро распространяться в воде. Весьма существенно, что у всех гидрофильных растений пыльца при попадании в воду не лопается (что происходит с пыльцой наземных растений) и остается жизнеспособной. Гидрофилия по сравнению с анемофилией довольно редкий тип абиотического перекрестного опыления.
Биотическое опыление может осуществляться как различными насекомыми (энтомофилия) — пчелами, шмелями, дневными и ночными бабочками, осами, жуками, перепончатокрылыми, муравьями, так и позвоночными животными — птицами, летучими мышами, мелкими приматами. В процессе длительной эволюции у растений выработались определенные приспособления для привлечения беспозвоночных и позвоночных животных-опылителей. Такими приспособлениями являются цветки с крупным околоцветником, определенная окраска венчика, наличие специфических запахов, характерная форма цветков, часто имитирующая форму тела самок насекомых-опылителей, большое количество пыльцы, присутствие нектара и т. д. Посещение опылителями цветков различного типа связано с их пищевыми потребностями в различные периоды жизненного цикла.
Среди многих исследователей, к сожалению, распространено представление о самоопылении как о тупике эволюционного развития. Оно сложилось под влиянием идей Ч. Дарвина, который показал приспособления цветков покрытосеменных растений к перекрестному опылению и описал те из них, которые препятствуют самоопылению.
У перекрестноопыляемых растений действительно существуют специальные приспособления, препятствующие самоопылению. К ним относятся дихогамия и гетеростилия. При дихогамии наблюдается неодновременное созревание тычинок и пестиков, проявляется она в форме протерандрии (протандрия) (более раннее созревание тычинок) (рис. 189) и протерогинии (протогиния) (более раннее созревание пестиков) (рис. 190).
Рис. 189. Явление протерандрии у камнеломки жестколистной (Saxifraga aizoides): А—цветок с хорошо развитыми тычинками и недоразвитыми стилодиями; Б—цветок с хорошо развитыми стилодиями и засыхающими пыльниками тычинок

Рис. 190. Явление протерогинии у норичника узловатого (Scrophularia nodosa) (продольный разрез цветка): А—цветок с хорошо развитым столбиком и недоразвитыми тычинками; Б—цветок с хорошо развитыми тычинками и увядающим столбиком

Протерандрия встречается чаще, чем протерогиния, и характерна для представителей семейств Зонтичные, Колокольчиковые, Кипрейные, Гераниевые и ряда других. Протерогиния наблюдается у многих крестоцветных, подорожниковых, у некоторых сложноцветных (дурнишник — Xanthium). При дихогамии разница во времени созревания тычинок и пестиков у разных видов варьирует от нескольких часов до нескольких дней.
Гетеростилия — особый тип приспособлений, который встречается у представителей семейств Первоцветные, Гречишные, Гвоздичные, Бурачниковые. При гетеростилии в разных цветках одного и того же вида образуются тычинки и столбики пестиков, имеющие различную длину. В цветках с длинными тычинками пестики имеют короткий столбик (или короткие стилодии). В цветках с короткими тычинками столбик пестика длинный (или длинные стилодии). Оплодотворение происходит только в том случае, если пыльца цветков с длинными тычинками попадет на рыльце пестика, имеющего длинный столбик, или если пыльца с цветков с короткими тычинками попадает на рыльце пестика с коротким столбиком. В других случаях пыльца не прорастает, что связано с самонесовместимостью пыльцы и рыльца одного цветка (рис. 191).
Рис. 191. Явление гетеростилии у гречихи посевной (Fagopyrum sagittatum): А—цветок с длинными стилодиями и короткими тычинками; Б—цветок с короткими стилодиями и длинными тычинками: 1—стилодии; 2—тычинки; 3—нектарники

В настоящее время бесспорно установлено, что в процессе эволюции у покрытосеменных растений выработалось два способа опыления — перекрестное и самоопыление. Несмотря на детерминированность способов опыления, они весьма лабильны, подвижны и могут в зависимости от климатических условий в период цветения меняться, т. е. у самоопылителей может наблюдаться перекрестное опыление, а у перекрестноопыляемых растений — самоопыление. Эволюционное значение такой подвижности имеет большое значение. При свободном переопылении внутри больших популяций очень затруднено закрепление новых комбинаций скрещивания. Любая удачная комбинация скрещивания при свободном переопылении будет распадаться и не сможет удержаться в потомстве. Самоопыление у перекрестников способствует изоляции новых форм, закреплению их в последующих поколениях, а значит, способствует видообразованию. Перекрестное опыление, которое время от времени наблюдается у самоопылителей, повышает уровень гетерозиготности потомства, а, следовательно, и жизнеспособность вида.
8.1.1. Прорастание пыльцы на рыльце пестика
Пыльца, попав тем или иным способом на рыльце пестика, прикрепляется к нему и начинает прорастать. Прорастанию пыльцы на рыльце пестика предшествует ряд физиолого-биохимических и цитологических процессов: выделение липидных капель на поверхности пыльцевого зерна, поступление воды из тканей пестика в пыльцу, активизация ферментов, перераспределение в цитоплазме всех компонентов пыльцевого зерна. Первым признаком того, что пыльца начала прорастать, т. е. формировать пыльцевую трубку, является циклоз — круговое вращательное движение цитоплазмы пыльцевого зерна. При прорастании пыльцы интина выпячивается через одну из апертур и образует пыльцевую трубку, в которую постепенно перемещается все содержимое пыльцевого зерна: цитоплазма вегетативной клетки с ядром, всеми органоидами и генеративной клеткой (если пыльца двуклеточная) или двумя спермиями (если пыльца трехклеточная) (рис. 192).
Рис. 192. Прорастающее пыльцевое зерно (пыльца) бобов обыкновенных (Vicia faba): 1—пыльцевое зерно; 2—пыльцевая трубка; 3—ядро вегетативной клетки; 4— генеративная клетка

Пыльцевая трубка внедряется в рыльце, затем проникает в столбик и начинает по нему продвигаться в сторону завязи. Столбик имеет различное строение. Он может быть открытым, закрытым, полуоткрытым. В зависимости от этого пыльцевая трубка продвигается по столбику по-разному. В открытом столбике она скользит по стенке канала. В закрытом и полуоткрытом пыльцевая трубка проходит, разрушая выстилающие столбик железистые клетки, и переходит на стенку завязи. Далее пыльцевая трубка движется по стенке завязи, пока не приблизится к месту расположения семяпочки.
Пыльцевая трубка растет по градиенту концентрации ростовых веществ, а также ионов кальция. Однако, вполне вероятно, что существуют и иные хемотропические факторы, которые способствуют продвижению пыльцевой трубки к семяпочке. Так как при продвижении по столбику и стенкам завязи пыльцевая трубка тесно контактирует с тканями материнского организма, очевидно, что между ними устанавливается какое-то специфическое физиологическое взаимодействие, которое пока еще не изучено.
У двудольных растений, для которых характерна двуклеточная пыльца, в пыльцевой трубке генеративная клетка делится митотически и образует два спермия (рис. 193). Если пыльцевая трубка широкая, митоз происходит типично, при митотическом делении ядра генеративной клетки четко вырисовываются все стадии митоза: профаза, метафаза, анафаза, телофаза. В узких пыльцевых трубках не образуется типичной метафазной пластинки. За митотическим делением ядра следует цитокинез. Таким образом, спермии являются гаплоидными клетками, имеющими, кроме плазмалеммы, клеточные оболочки. Цитоплазма спермиев занимает в клетке боковое положение, а также сконцентрирована на полюсах. Ядро не имеет ядрышка, а хроматин ядра находится в конденсированном состоянии. В типичном случае в цитоплазме спермиев содержится характерный для растительных клеток набор органоидов: митохондрии, диктиосоммы, гранулярный эндоплазматический ретикулум, рибосомы, микротрубочки. Имеются также небольшие вакуоли и пластиды. У некоторых злаков, сложноцветных отмечен диморфизм спермиев, который проявляется не только в различии формы, размеров, но и в содержании органелл.
Рис. 193. Спермий в пыльцевой трубке бобов обыкновенных (Vicia faba)

Вопрос о движении генеративной клетки и спермиев в пыльцевой трубке является проблематичным, а представления различных авторов о способах движения — противоречивыми. Одни авторы считают, что генеративная клетка и спермии, независимо от места их образования, движутся активно, другие указывают, что они передвигаются пассивно, увлекаемые током цитоплазмы. Не ясна и функция вегетативного ядра. Некоторые авторы на основании того, что оно всегда располагается в растущем конце пыльцевой трубки, считают, что вегетативное ядро регулирует рост пыльцевой трубки, которая обладает специфической поляризацией. По мере того как пыльцевая трубка удлиняется, пыльцевое зерно (пыльца) и основание пыльцевой трубки опустошаются, и живое содержимое отделяется от пустого участка каллозной пробкой. Таким образом сохраняется целостность мужского гаметофита. Для пыльцевой трубки характерен только апикальный рост, растущая зона отделяется от остальной части пыльцевой трубки все новыми и новыми каллозными пробками.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Органом полового размножения покрытосеменных растений является цветок. Цветок — видоизмененный, укороченный, неразветвленный побег, предназначенный для образования спор и гамет и полового процесса, завершающегося образованием семян и плода.
Строение цветка
У цветка различают цветоножку, цветоложе, околоцветник, тычинки и пестики. У некоторых цветков отдельные части могут отсутствовать.
Цветки большинства видов растений имеют и тычинки, и пестики. Такие цветки называют обоеполыми (вишня, горох). Цветки, которые имеют только пестики, называют пестичными (женскими). Цветки, которые имеют только тычинки, называют тычиночными (мужскими). В зависимости от распределения однополых цветков на растениях различают: однодомные растения — растения, у которых на одних и тех же экземплярах располагаются и женские, и мужские цветки (огурец, кукуруза, дуб); двудомные растения — растения, у которых на одних экземплярах располагаются женские, а на других — мужские цветки (крапива двудомная, конопля, облепиха); многодомные растения — растения, у которых на одних и тех же экземплярах встречаются как обоеполые, так и однополые цветки в различных количественных соотношениях (гречиха, некоторые виды ясеня, клена).
Цветоножка — междоузлие под цветком. Цветки, лишенные цветоножки, называются сидячими (цветки в соцветии корзинка у подсолнечника, астры, одуванчика).
Цветоложе — укороченная стеблевая часть цветка. На ней располагаются все остальные части цветка.
Околоцветник — стерильная часть цветка, его покров. Околоцветник может быть простым (не дифференцированным на чашечку и венчик, образованным совокупностью однородных листочков, имеющих одинаковые размеры и окраску) и двойным (дифференцированным на чашечку и венчик, отличающиеся друг от друга размерами и окраской. Простой околоцветник может быть венчиковидным (образованным ярко окрашенными листочками) или чашечковидным (образованным зелеными листочками). Цветки, лишенные околоцветника (ива, тополь), называются голыми.
Чашечка — наружная часть двойного околоцветника, представляет собой совокупность чашелистиков — видоизмененных прицветных листьев. Обычно чашелистики имеют небольшие размеры и зеленую окраску. Они сходны с обычными листьями, но устроены проще.
Различают: раздельнолистную чашечку — чашечку, образованную свободными (несросшимися) чашелистиками (капуста, лютик); сростнолистную чашечку — чашечку, образованную частично или полностью сросшимися чашелистиками (картофель, табак, горох).
Венчик — внутренняя, обычно окрашенная часть двойного околоцветника. Представляет собой совокупность лепестков, часто имеющих яркую окраску. Количество лепестков венчика может быть различным. Лепестки могут быть более или менее одинаковыми (лютик,яблоня) либо отличаться размерами и формой (фиалка, горох). В результате венчик может быть правильным, неправильным или асимметричным. Венчик, как и чашечка, может быть раздельнолепестным и сростнолепестным. Раздельнолепестной венчик состоит из свободных, несросшихся лепестков. Сростнолепестной венчик состоит из сросшихся в той или иной степени лепестков. Главная функция венчика — привлечение опылителей.
Андроцей
Андроцей — совокупность тычинок одного цветка. Количество тычинок в цветке — от одной (орхидные) до нескольких сотен (некоторые кактусы). У большинства растений тычинка состоит из тычиночной нити и пыльника. Тычиночная нить — нижняя, как правило, суженная стерильная часть тычинки. Нижний конец тычиночной нити отходит от цветоложа, а верхний конец несет пыльник. Обычно тычиночные нити тонкие, длинные, округлые в сечении. Пыльник — верхняя расширенная фертильная часть тычинки. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда (микроспорангия), в которых происходит образование микроспор, а впоследствии пылинок. Связник является продолжением тычиночной нити, через него в пыльник поступают питательные вещества.
Микроспорогенез
Микроспорогенез — процесс образования микроспор в микроспорангиях (гнездах пыльника). Микроспоры формируются из материнских клеток — микроспороцитов, имеющих диплоидный набор хромосом. В результате мейоза каждая материнская клетка образует четыре гаплоидных микроспоры. Микроспоры быстро обособляются друг от друга.
Микрогаметогенез
Микрогаметогенез — процесс образования мужских половых клеток (спермиев), происходит в пыльцевом зерне, которое является мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита происходит также в гнездах пыльников тычинок и сводится к одному митотическому делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины (наружной, толстой). Каждое пыльцевое зерно содержит две гаплоидные клетки: вегетативную и генеративную. Из генеративной (спермагенной) далее образуются два спермия. Из вегетативной (сифоногенной) впоследствии образуется пыльцевая трубка.
Гинецей
Гинецей — совокупность пестиков одного цветка. Обычно в пестике выделяют три части: завязь, столбик и рыльце.
Завязь — замкнутая, нижняя, полая часть пестика, несущая и защищающая семязачатки. Завязь бывает: верхняя, нижняя, полунижняя. В завязи может располагаться от одного (пшеница, вишня) до нескольких тысяч (мак) семязачатков. Стенки завязи выполняют функцию защиты семязачатков от неблагоприятных факторов среды (высыхание, колебание температур, поедание насекомыми и т.д.). Внутри завязи (в семязачатках) происходит мегаспорогенез и мегагаметогенез, они принимают участие в образовании околоплодника.
Столбик — средняя более или менее удлиненная стерильная часть пестика, отходящая обычно от верхушки завязи, соединяет завязь и рыльце.
Рыльце — верхняя расширенная часть пестика, предназначено для восприятия пыльцы. Рыльце может быть разнообразной формы (двухлопастное, звездчатое, перистое и т.д.) и размера в зависимости от особенностей опыления. При отсутствии столбика рыльце называют сидячим.


Семязачаток состоит из нуцеллуса (ядра) — центральной части, являющейся мегаспорангием, двух покровов — интегументов, которые при смыкании образуют узкий канал — микропиле, или пыльцевход, через который пыльцевая трубка проникает к зародышевому мешку. С помощью семяножки семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где сливаются нуцеллус и интегументы, называют халазой.
В семязачатке происходят мегаспорогенез, мегагаметогенез и процесс оплодотворения. После оплодотворения (реже без него) из семязачатка формируется семя.

Мегаспорогенез
Процесс формирования мегаспор называется мегаспорогенезом. Он происходит в нуцеллусе семязачатка. После заложения семязачатка и формирования нуцеллуса в области микропиле начинает разрастаться одна археспориальная (спорогенная) клетка — мегаспороцит, или материнская клетка мегаспор.
Материнская клетка мегаспор имеет диплоидный набор хромосом. У большинства покрытосеменных из нее путем мейоза образуются четыре гаплоидные мегаспоры. Из них лишь одна (обычно нижняя, обращенная к халазе, реже верхняя, обращенная к микропиле) дает начало женскому гаметофиту — зародышевому мешку. Остальные мегаспоры отмирают.
Мегагаметогенез
Процесс формирования женских половых клеток происходит в зародышевом мешке. Формирование женского гаметофита начинается с разрастания мегаспоры, которая далее три раза делится митозом. В результате этого образуются восемь клеток, которые располагаются следующим образом: три — на одном полюсе зародышевого мешка (микропилярном), три — на другом (хадазальном), две — в центре. Две оставшиеся сливаются в центре клетки, образуя диплоидную центральную клетку зародышевого мешка. Одна из трех клеток, расположенных на микропилярном полюсе, отличается большими размерами и является яйцеклеткой. Две рядом расположенные клетки являются вспомогательными и называются синергидами. Группа из трех клеток, находящихся на противоположном, халазальном полюсе, называется антиподом. Таким образом, сформированный женский гаметофит включает шесть гаплоидных клеток (яйцеклетка, две клетки-синергиды, три клетки-антипода) и одну диплоидную клетку.
Оплодотворение. Образование семян и плодов
Купить проверочные работы
и тесты по биологии




Процессу оплодотворения предшествует опыление — перенос пыльцы от пыльцевых мешков тычинок к рыльцам пестиков. Попав на рыльце пестика, под воздействием веществ, выделяемых пестиком, пыльца начинает прорастать: образуется пыльцевая трубка, внедряющаяся в ткань рыльца. Кончик пыльцевой трубки выделяет вещества, размягчающие ткань рыльца и столбика. В процессе формирования пыльцевой трубки принимает участие сифоногенная клетка. По мере роста пыльцевой трубки в нее переходит спермагенная клетка, которая делится митозом с образованием двух спермиев (у некоторых растений спермагенная клетка дает начало двум спермиям еще до прорастания пыльцы). Пыльцевая трубка продвигается по столбику пестика и врастает в зародышевый мешок, как правило, через микропиле. После проникновения в зародышевый мешок кончик пыльцевой трубки разрывается, и спермии попадают внутрь. Один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу. Второй спермий сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку, из которой далее формируется эндосперм (питательная ткань) семени, обеспечивающий питание зародыша. Синергиды и антиподы дегенерируют. Вышеописанный процесс получил название двойного оплодотворения. Двойное оплодотворение у цветковых растений было открыто в 1898 году русским ботаником С.Г. Навашиным.
После двойного оплодотворения из яйцеклетки формируется зародыш семени, из центрального ядра зародышевого мешка — эндосперм, из интегументов — семенная кожура, из всего семязачатка — семя, а из стенок завязи — околоплодник. В целом из завязи пестика формируется плод с семенами.

Общая характеристика. Моховидные — это наиболее примитивная группа высших растений с особой линией развития. Подразделяются на два класса (печеночные и листостебельные мхи) и насчитывают около 20 тыс. видов. Широко распространены во всех широтах от тундры до тропиков. Большинство видов — тенелюбивые. Растут на почве, на стволах растений, на скалах и стенах домов. Некоторые виды живут в воде.
Главная отличительная особенность: в жизненном цикле моховидных преобладает гаметофит, а спорофит редуцирован и развивается на гаметофите.
Прогрессивный признак моховидных — наличие дифференцированных тканей: эпидермиса, основной (ассимиляционной паренхимы), проводящего пучка. Механическая ткань отсутствует.
Строение. Моховидные представляют собой небольшие (до нескольких сантиметров), многолетние (редко однолетние) растения. Тело печеночных мхов представлено слоевищем. У листостебельных мхов тело гаметофита состоит из простых или разветвленных стеблей, покрытых мелкими листьями с одной жилкой. Листостебельный побег покрыт эпидермисом, под которым располагается кора; в центре находится проводящий пучок, состоящий из удлиненных мертвых клеток (они проводят воду и минеральные вещества) и окружающих их живых клеток, проводящих органические вещества.
Корни отсутствуют, их роль выполняют вытянутые клетки в нижней части стебля (ризоиды). Настоящих сосудов нет. На гаметофите вместо устьиц имеются поры, лишенные замыкающих клеток.
У сфагновых мхов стебель гаметофита несет мутовки ветвей, густо покрытых листьями. Листья не имеют средней жилки. Клетки в листьях дифференцированы на ассимиляционные (с хлоропластами) и мертвые водосборные клетки с порами в клеточной оболочке. Ризоиды отсутствуют; воду и минеральные вещества эти мхи всасывают стеблем и листьями, являясь мощным сорбентом воды, и выделяют в почву гуминовые кислоты, обладающие бактерицидным действием. Нарастают верхушкой, нижняя часть стебля отмирает, но полностью не сгнивает, образуя торф.
Размножение. В жизненном цикле происходит правильное чередование полового и бесполого поколений. Половое поколение (гаметофит с гаплоидным набором хромосом) представлено зелеными растениями и преобладает над бесполым (спорофитом). На гаметофите образуются органы полового размножения — антеридии и архегонии. В антеридиях созревают подвижные двужгутиковые споры, а в архегониях — по одной неподвижной яйцеклетке. Оплодотворение происходит в капельножидкой среде. После слияния гамет образуется зигота, которая дает начало бесполому поколению — спорофиту. Спорофит развивается на гаметофите и представляет собой коробочку, расположенную на ножке и прикрываемую колпачком (калиптрой). Ножка спорофита в нижней части переходит в стопу, с помощью которой он прикрепляется к телу гаметофита. Бесполое размножение осуществляется спорами. Образование спор на спорофите происходит в спорангиях, расположенных внутри коробочек. Споры гаплоидны. При попадании в благоприятную среду они прорастают с образованием разветвленной нити (протонемы). На ней закладываются почки, из которых прорастают листостебельные побеги мха.
У мхов широко распространено вегетативное размножение: почками, молодыми побегами, листьями, выводковыми телами.

Значение. Мхи одними из первых заселяют бесплодные участки (камни, скалы, пески). При отмирании создают обогащенный органическими веществами субстрат, пригодный для поселения других высших растений. Формируют в лесу покров (ковер), способствующий поддержанию водного баланса почвы и обеспечивающий постепенный переход поверхностного стока воды в подземный, предотвращая эрозию почвы. Сфагновые мхи могут приводить к заболачиванию местности и принимают участие в образовании торфа (используется как удобрение, топливо, сырье).
Папоротниковидные
Общая характеристика. Папоротниковидные — высшие споровые, преимущественно травянистые растения с большими раскидистыми листьями, стеблями и корнями; реже встречаются древесные формы. Произошли в девонский период палеозойской эры от риниофитов. В настоящее время сохранилось около 12 тыс. видов. Широко распространены во всех частях света; наиболее многочисленны в Юго-Восточной Азии. Обитают в тенистых и сырых местах.
Основная жизненная форма — диплоидный спорофит (листостебельное растение).
Размножение. В жизненном цикле происходит правильное чередование полового и бесполого поколений. Половое поколение (гаметофит) представлено заростком, бесполое поколение (спорофит) — самим растением со спорангиями на листьях. Спорофит преобладает над гаметофитом. В спорангиях образуются споры, которые при созревании разносятся ветром и, попав на влажную почву, прорастают, образуя гаметофит (заросток). Гаме-тофит представляет собой гаплоидную зеленую пластинку размером около 0,5 см, на которой формируются архегонии и антеридии. В архегониях созревают многожгутиковые сперматозоиды, в антеридиях — неподвижные яйцеклетки. Оплодотворение происходит в капельно-жидкой среде с образованием зиготы. Из зиготы образуется зародыш, из которого прорастает взрослое растение.

Значение. В умеренном климате роль папоротников незначительна. В странах с теплым влажным климатом папоротники -важный компонент многих растительных сообществ. Человеком часто используются как декоративные растения (асплениум, нефролепис и др.). Некоторые виды папоротников употребляются в пищу, (орляк); их экстракты применяются для лечения болезней легких, изгнания глистов.
Плауновидные
Плауны — высшие споровые многолетние травянистые лесные растения размером от нескольких десятков сантиметров до 3 м, внешне напоминающие некоторые виды мхов. Насчитывают около 400 видов. Основная жизненная форма — диплоидный спорофит.
Особенности строения. Плауны имеют стелющиеся и приподнимающиеся, дихотомически ветвящиеся побеги. Листья простые, цельные, мелкие, сидячие, линейно-шиловидные, располагаются поочередно, иногда дихотомически.
Размножаются спорами, образующимися в результате мейоза и содержащих гаплоидный набор хромосом. Споры созревают в спороносных колосках — стробилах, образующихся на верхушках побегов и имеющих длину 15-20 см. Споры имеют тетраэдрическую форму и две оболочки — внутреннюю и наружную (шиповатую). Из споры прорастает гаметофит — заросток. Живет он частично или полностью под землей до 15 и более лет, питаясь благодаря симбиозу с грибами. После его появления над землей на нем формируются антеридии и архегонии. При наличии капельно-жидкой среды сперматозоид из антеридия проникает в архего-ний, где происходит оплодотворение. Образуется зигота, из которой прорастает зародыш спорофита, который после укоренения начинает существовать самостоятельно.
Значение. Споры плаунов используются в медицине при лечении пролежней и в качестве детской присыпки. Древовидные формы, существовавшие в карбоне, внесли значительный вклад в формирование залежей каменного угля.
Хвощевидные
Хвощи — это высшие споровые многолетние травянистые лесные растения размером 40-60 см (некоторые виды — до 10-12 м). Были широко распространены в палеозое. В настоящее время представлены одним родом — хвощом, который насчитывает около 20 видов. Распространены везде, кроме Австралии и Новой Зеландии. Растут на полях, лугах, болотах, по берегам водоемов, реже на сухих песчаных почвах в сосновых лесах. Жизненный цикл осуществляется с чередованием поколений и подобен жизненному циклу папоротников. Основная жизненная форма — диплоидный спорофит.
Особенности строения. Хвощи имеют членистое, ветвистое корневище с придаточными корнями. Надземные побеги — обычно двух видов: вегетативные (зеленые, летние) и спороносные (бурые, неветвящиеся, весенние, несущие на верхушке спороносный колосок). Для хвощей характерно членение стебля на четко выраженные ребристые междоузлия и вздутые узлы, к которым прикрепляются мутовки боковых побегов. Чешуевидные листья также располагаются мутовчато, срастаясь между собой в трубку почти до самой верхушки. Листья не способны к фотосинтезу, эту функцию выполняет зеленый стебель и побеги, имеющие хорошо развитую ассимиляционную ткань, сосудисто-волокнистые проводящие пучки и содержащие участки механической ткани.
Размножение. Хвощи размножаются при помощи спор, образующихся в специальных колосках, развивающихся либо на спороносных, либо на обычных побегах. Из спор после их падения на землю вырастают заростки (половое поколение), на которых затем формируются архегонии и антеридии, где созревают гаметы. Оплодотворение происходит в капельно-жидкой среде. После него образуется зигота, затем формируется спора, из которой прорастает взрослое растение — спорофит.
Значение. Хвощ полевой используется как лекарственное растение в качестве мочегонного и кровеостанавливающего средства. Некоторые виды хвоща используются для чистки посуды и мебели. Хвощ болотный ядовит для животных, хвощ полевой является трудноискоренимым сорняком.
Голосеменные
Общая характеристика. Голосеменные — это семенные растения, имеющие незащищенные семяпочки, расположенные открыто на семенной чешуе шишки; семена защищены только семенной кожурой. Голосеменные не имеют настоящих цветков и не образуют плодов. Они являются наиболее древними из семенных растений; произошли от семенных папоротников в начале мезозойской эры (около 350 млн. лет назад). В настоящее время насчитывают около 650 видов.
В жизненном цикле происходит правильное чередование полового и бесполого поколений. Основная жизненная форма — диплом идный спорофит (само растение), гаметофит редуцирован, утратил самостоятельность и живет на спорофите. Мужской гаметофит не имеет антеридиев. Растения опыляются в основном ветром.
Особенности строения. Спорофит голосеменных — это одно-или двудомное растение с хорошо развитыми стеблем (имеющим моноподиальное ветвление) и стержневой корневой системой с выраженными главным и боковыми корнями. Представлен в основном древесными вечнозелеными формами, реже встречаются листопадные деревья (лиственница), имеются кустарники (можжевельник) и лианы (эфедра); травянистых растений нет. Листья игловидные (хвоя) или чешуевидные; располагаются поодиночке, по два или несколько в пучках. В древесине большинства голосеменных нет настоящих сосудов и древесных волокон; трахеиды выполняют водопроводящую и опорную (механическую) функции.
Важнейшие ароморфозы: появление семени и пыльцевой трубки, что обеспечило опыление и оплодотворение без воды.
Характерные особенности образования семян: оплодотворение — простое (в нем участвует один спермий из двух, имеющихся в пыльцевом зерне), эндосперм образуется в семязачатке до оплодотворения, а семена покрыты только семенной кожурой.
Голосеменные широко распространены во всех частях света; составляют большинство и доминируют в растительном покрове в умеренном климатическом поясе. Подразделяются на четыре класса: Хвойные, Гнетовые, Саговниковые, Гинкговые.
Класс Хвойные — вечнозеленые растения, имеющие видоизмененные игольчатые листья. Листья игловидные, покрыты кутикулой или восковым налетом; устьиц мало, глубоко посажены (это обеспечивает жизнестойкость растений в засушливые и зимние периоды). У хвойных стебель прямостоячий, покрыт корой, содержащей ситовидные клетки, механическую ткань, паренхиму, смоляные ходы, камбий. Древесина на 90-95% состоит из трахеид и не содержит механической ткани. Трахеиды имеют плотные од-ревесневевшие стенки. Сердцевина представлена основной тканью.
Сосна обыкновенная — это однодомное светолюбивое растение, у которого весной формируется раздельнополые мужские и женские шишки (стробилы). Мужские шишки формируются у основания побега, имеют зеленовато-желтый цвет и длину около 5 мм. На осях таких шишек расположены слои мелких чешуек -микроспорофилл. На нижней поверхности каждой из чешуек расположены два микроспорангия (пыльцевых мешка), в которых образуется пыльца (мужской гаметофит). Каждое пыльцевое зерно содержит вегетативную клетку, два спермия и имеет два воздушных мешка, облегчающих перенос пыльцы ветром,
Женские шишки имеют красноватый цвет и содержат крупную, толстую семенную чешую (с двумя семязачатками на поверхности каждой чешуйки) и мелкую прозрачную кроющую чешую. В каждом семязачатке формируется женский гаметофит, который содержит гаплоидный эндосперм (особую питательную ткань) и два архегония с крупной яйцеклеткой в каждом. Семязачаток снаружи покрыт интегументом и имеет пыльцевход (микропиле), через который пыльца втягивается внутрь, где прорастает с формированием пыльцевой трубки. По ней созревшие спер-мии проникают к архегониям. Оплодотворение простое и не требует присутствия воды; в нем участвует только один из спермиев, сливающийся с одной из яйцеклеток, второй спермий погибает. Образовавшаяся зигота делится, из нее формируется зародыш семени, а вся семяпочка превращается в семя.
У сосны обыкновенной семена созревают на второй год, разносятся ветром и в благоприятных условиях прорастают.


Знамение голосеменных: выделяют в атмосферу кислород и фитонциды, являются источником сырья для строительной, деревообрабатывающей и целлюлозной промышленности, получения скипидара, канифоли, спирта, лекарств; многие используются в декоративных целях (ели, туя, саговниковые); семенами многих хвойных питаются птицы (клесты, синицы, дятлы) и млекопитающие; семена кедровой сосны и пинии используются в пищу человеком.
Покрытосеменные (цветковые)
Общая характеристика. Покрытосеменные — самый большой отдел растительного мира. Произошли от древней формы голосеменных в начале мелового периода мезозойской эры (около 125 млн. лет назад). Насчитывают около 250 тыс. видов. Занимают господствующее положение в растительном мире; произрастают во всех климатических зонах и в самых разных климатических условиях. Наибольшее разнообразие видов — во влажных тропиках.
Покрытосеменные — единственная группа растений, образующая сложные многоярусные фитоценозы.
Особенности строения. Доминирующая жизненная форма -диплоидный спорофит (само растение, включающее корень и побег). Спорофиты разных групп покрытосеменных представлены различными жизненными формами (древесными, кустарниковыми, кустарничковыми, лиановыми, одно- и многолетними травами) и могут очень сильно отличаться друг от друга. Большинство покрытосеменных — многолетние растения, а травы могут быть одно- и двулетними. Органы покрытосеменных делятся на вегетативные (корень, стебель, лист) и генеративные (цветок, плод, семя).
В зависимости от строения семян и морфологических особенностей органов отдел делится на два класса: Однодольные (зародыш семени имеет одну семядолю) и Двудольные (зародыш семени имеет две семядоли). Основные различия между одно- и двудольными растениями приведены в таблицах на с. 280-281. При делении классов на семейства, роды и виды учитываются общие признаки растений — строение цветка и плода, тип соцветия, особенности внешнего и внутреннего строения вегетативных органов.
Основные ароморфозы: цветок, плод, двойное оплодотворение; симподиальное ветвление, прогрессивное развитие проводящей ткани: ксилема содержит настоящие сосуды — широкие трахеи (а не трахеиды, как у голосеменных), флоэма — ситовидные трубки с клетками-спутницами, а не ситовидные клетки; наличие специализированной механической ткани (волокон), придающей прочность коре и древесине; прогрессивное развитие вегетативных органов; способность образовывать ядовитые вещества, защищающие растения от растительноядных животных (у некоторых групп покрытосеменных).
Семязачатки находятся в полости завязи пестика и защищены его тканями от неблагоприятных условий среды, семена защищены не только семенной кожурой, но и плодом.
Развитие эндосперма одновременно с развитием зародыща (а не до оплодотворения, как у голосеменных) позволяет избежать ненужной траты питательных веществ и энергии в том случае, когда зародыш не образуется.
Гаметофиты более упрощены, чем у голосеменных. Мужской гаметофит покрытосеменных представлен пыльцевым зерном, содержащим вегетативную клетку с диплоидным набором хромосом (2n) и генеративную гаплоидную клетку (1n). У некоторых до опыления путем митоза из генеративной клетки образуется два спермия с гаплоидным набором хромосом. Женский гаметофит представлен зародышевым мешком, содержащим две клетки -синергиды, три клетки-антиподы, яйцеклетку с гаплоидным набором хромосом (1n) и диплоидную центральную клетку (2n).
Размножение. Пыльцевые зерна попадают на рыльце пестика (а не сразу в пыльцевход семязачатка), предназначенного именно для улавливания пыльцы; в пыльцевход семязачатка проникает пыльцевая трубка, образующаяся в пыльцевом зерне. Оплодотворение двойное: в нем участвуют два спермия: один сливается с яйцеклеткой, образуя диплоидную зиготу, из которой развивается зародыш, другой спермий сливается с диплоидной центральной клеткой женского гаметофита, образуя триплоидную клетку, из которой впоследствии возникает эндосперм, содержащий запас питательных веществ, необходимых для развития зародыша.
После оплодотворения семязачаток развивается в семя, завязь пестика формирует плод. Наличие и уникальность плода обеспечивают распространение покрытосеменных птицами, млекопитающими, насекомыми, ветром, водой и т.д.
Для покрытосеменных также характерно вегетативное размножение (с помощью вегетативных органов).

Значение покрытосеменных: поддерживают стабильный газовый состав атмосферы; образуют сложные многоярусные фитоценозы; растения и их плоды служат пищей для многих видов животных; покрытосеменные широко используются человеком: обеспечивают человека хлебом (хлебные злаки), используются им в пищу (овощные и плодово-ягодные) и в качестве лечебных средств, являются источником сырья для деревообрабатывающей, целлюлозной, легкой и медицинской промышленности, применяются в декоративных целях и т.д.

В видеоуроке рассматриваются отделы Голосеменные и Покрытосеменные растения, особенности строения их органов размножения, а также особенности размножения голосеменных и покрытосеменных и процесс двойного оплодотворения цветковых. В данном уроке приводятся следующие понятия: семязачаток, оплодотворение, репродуктивные органы, генеративные органы, завязь, семяпочка, двойное оплодотворение, опыление.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Семенные растения. Отдел Голосеменные и Покрытосеменные. Оплодотворение у цветковых растений"
Все растения можно разделить на низшие и высшие. Высшие растения подразделяют на споровые и семенные. А высшие семенные растения подразделяют на голосеменные и покрытосеменные (цветковые).
Голосеменные — это исключительно наземные вечнозелёные, реже листопадные деревья.
Своё название голосеменные получили потому, что их семена лежат открыто на поверхности чешуи шишек.
Наличие семян создаёт им огромное преимущество перед споровыми.
Из современных голосеменных наиболее известны хвойные. К ним относятся ель, сосна, пихта, кедр, можжевельник, лиственница и др.
Голосеменные имеют стебель, корень, листья.
У большинства представителей листья представлены хвоей. Хвоя имеет плотную кожицу, покрытую восковидным веществом, поэтому растения испаряют мало воды и хорошо приспособлены к неблагоприятным условиям.
Отдел Голосеменные включает четыре класса: класс Гинкговые, класс Гнетовые, класс Саговниковые (Цикадовые), класс Хвойные
Наиболее широко распространёнными представителями хвойных в России являются сосна обыкновенная и ель обыкновенная, или европейская.
Сосна, как и все хвойные растения, размножается семенами. Она светолюбива, поэтому в сухих сосновых лесах (борах) всегда светло.
В мае у основания молодых побегов сосны образуются пучки зеленовато-жёлтых мужских шишек длиной 4—6 мм.
На других побегах того же растения образуются женские шишки. Сначала они зелёные, а потом становятся красновато-коричневыми.

Каждая шишка состоит из оси и сидящих на ней чешуй.
На чешуях мужских шишек развивается по два пыльцевых мешочка. В них созревает пыльца. Оболочка каждой пылинки имеет два пузырька, наполненных воздухом. Такие пылинки ветер может переносить на большие расстояния.
На чешуе женских шишек развивается по два семязачатка, в которых находятся женские гаметы (яйцеклетки).
Если пыльца с мужских шишек попадёт на женские шишки, произойдёт опыление. После этого чешуи женских шишек смыкаются и склеиваются смолой.
Оплодотворение, то есть слияние спермия и яйцеклетки, у сосны происходит через год после опыления.
Пыльца с одного дерева при помощи ветра перелетает на другое и, попав на женскую шишку, прорастает в пыльцевую трубку, в которой образуются мужские гаметы — два спермия. Пыльцевая трубка достигает семязачатка, где находится яйцеклетка, затем происходит оплодотворение. Из зиготы развивается зародыш, а из всего семязачатка — семя.
Таким образом, семена в шишках сосны созревают через год после опыления.
Семя находится на поверхности чешуи женской шишки.
У большинства видов сосны семена имеют плёнчатые крылышки, благодаря которым могут распространяться.
Тем временем женские шишки растут и одревесневают, из зелёных они превращаются в коричневые.
Зрелое семя сосны выпадает из шишки и распространяется. В дальнейшем, попав в благоприятные условия, зародыш трогается в рост и начинает прорастать.
Всходы сосны имеют своеобразный вид. Это меленькие растеньица, у которых стебелёк очень тонкий. На верхушке стебелька находится пучок лучеобразно расходящихся во все стороны тонких иголочек-семядолей.
Покрытосеменные растения, как считают учёные, произошли от древних голосеменных. Это самая молодая и в то же время самая многочисленная группа царства Растения. Она включает примерно 250 тыс. видов, произрастающих во всех климатически зонах.
Покрытосеменные, или цветковые, — отдел наиболее совершенных высших растений, имеющих цветок. Цветок — это орган семенного размножения. Стеблевая часть цветка представлена цветоножкой, верхнюю часть которой называют цветоложем. Цветок снаружи окружает околоцветник. Он выполняет защитную функцию, и к тому же лепестки могут привлекать опылителей.
В середине цветка находятся репродуктивные части, участвующие в размножении: пестик (или пестики) и тычинки.
Пестик — это женский репродуктивный орган цветковых растений.
Тычинка — это мужской репродуктивный орган, в котором образуется пыльца.
Пыльца — порошкообразные споры, обычно жёлтого цвета, содержащие мужские половые клетки.

Цветок может иметь и пестик, и тычинки, и тогда он будет обоеполым цветком. У однополых цветков имеются или только тычинки (тычиночные, или мужские, цветки), или только пестик (пестичные, или женские, цветки).
Покрытосеменные растения, как и голосеменные, размножаются с помощью семян, которые образуются после отцветания. Но семена их защищены околоплодником, т. е. они покрыты, что способствует их лучшему сохранению и распространению.
Отсюда и название этого отдела высших растений ― покрытосеменные (или цветковые).
Все покрытосеменные, несмотря на своё многообразие, имеют общий план строения. Их органы подразделяют на репродуктивные, или генеративные, и вегетативные.
Репродуктивные, или генеративные, органы связаны с половым размножением растений. К ним относят цветок и плод с семенами.
Вегетативные органы составляют тело растения и осуществляют его основные функции, включая вегетативное размножение — это один из способов бесполого размножения. К вегетативным относят корень, стебель, лист.
Период жизни покрытосеменных растений может быть различным.
Если растение проживает только один вегетативный период (то есть период, когда возможен его рост и развитие), например с весны по осень, то такое растение называют однолетним. К однолетним растениям относят горох, кукурузу, астру и другие.
Двулетние растения в первый год накапливают питательные вещества в корнях и стеблях, а на следующий год заканчивают свою жизнь цветением и плодоношением (капуста, морковь, свёкла и другие).
У многолетних растений цветение и плодоношение может быть многократным. К таким растениям относят картофель, чеснок, лук и другие.
Размножение покрытосеменных растений связано с цветком.
Важные части цветка — это пестик и тычинка. С их участием происходят сложные процессы полового размножения — опыление и оплодотворение.
Но сперва в будущем цветке начинают образовываться половые клетки.
Женские половые клетки формируются в завязи пестика в семяпочке.
Одна из клеток семяпочки делится. В результате образуются 4 клетки — это женские споры цветкового растения. И только одна из четырёх клеток растёт. В ней содержится большое количество цитоплазмы.
Ядро этой клетки делится, в результате чего образуются 2 дочерних ядра. Ядра расходятся в противоположные стороны и вновь делятся дважды. В результате образуется восьмиядерная клетка. В верхних и нижних её частях образуется по четыре ядра. От каждой группы к центру перемещается по одному ядру. Вокруг остальных ядер образуется цитоплазма.
Одна из клеток становится женской гаметой — яйцеклеткой.
Ядра, содержащие гаплоидный набор хромосом, которые переместились к центру, сливаются, получается одно диплоидное вторичное ядро. Так образуется зародышевый мешок, который состоит из нескольких клеток. Это и есть гаметофит женского цветкового растения.
Посмотрим, как образуется мужской гаметофит.
Все клетки пыльника вначале однородны. Затем первичные спорогенные клетки начинают делится. В результате деления получаются гаплоидные микроспоры. Они находятся в гнёздах пыльника. Зрелая пыльцевая клетка (микроспора) одноядерна.
Каждая спора делится и образует две клетки: вегетативную и генеративную.
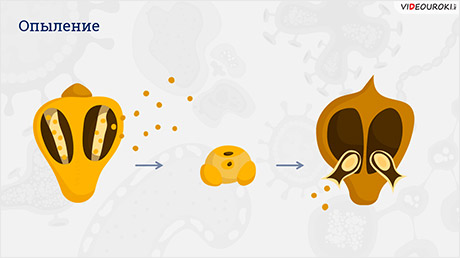
Пыльник созревает, и пыльца высыпается. Она несёт только генетическую информацию. Пылинка попадает на рыльце пестика и происходит опыление.
Под действием вещества, которое находится на рыльце, пыльца прорастает в направлении семяпочки. Из вегетативной клетки пыльцевого зерна образуется пыльцевая трубка.
Генеративная клетка делится, в результате образуются две мужские клетки гаметы ― спермии. А вегетативная клетка исчезает.
Пыльцевая трубка через пыльцевход прорастает в семязачаток. И обе мужские гаметы вливаются в зародышевый мешок.
Одна из гамет перемещается к яйцеклетке и сливается с ней.
Из двух гаплоидных гамет в результате оплодотворения возникает диплоидная зигота. Из неё развивается зародыш семени.
Вторая мужская гамета перемещается к диплоидному вторичному ядру, образуется ядро, которое содержит тройной набор хромосом.
Оплодотворение, при котором одна мужская гамета сливается с яйцеклеткой, а вторая ― с вторичным ядром, называется двойным оплодотворением.
Триплоидное ядро многократно делится. Из получившихся клеток образуется питательная ткань эндосперм.
К завязи притекают питательные вещества, и она постепенно превращается в спелый плод. Околоплодник защищает семена от неблагоприятных воздействий окружающей среды.
В последствии из него образуется новое растение. А богатые питательными веществами ткани эндосперма обеспечат проросток необходимым питанием.
Процесс двойного оплодотворения открыл в 1898 году русский ботаник, академик Сергей Гаврилович Навашин.
Прежде чем произойдёт оплодотворение, как мы уже говорили, должно произойти опыление. Различают два основных типа опыления: самоопыление и перекрёстное опыление.
При самоопылении пыльца из тычинки попадает на рыльце пестика того же самого цветка. Так опыляются пшеница, рис, горох и др.
При перекрёстном опылении пальца с тычинок цветка одного растения переносится на рыльца пестиков цветков других растений. В природе перекрёстное опыление происходит значительно чаще, чем самоопыление.
Читайте также: