
Двойное оплодотворение у растений реферат
Добавил пользователь Евгений Кузнецов Обновлено: 19.09.2024
половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка (См. Зародышевый мешок). Д. о. открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений — лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия (См. Спермии) сливается с ядром яйцеклетки, ядро второго — с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается Зародыш, из центральной клетки — Эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид (См. Синергиды), которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой — проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При Д. о. ядра зародышевого мешка находятся в интерфазе (См. Интерфаза) и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа Д. о.: премитотическое — ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое — мужское и женское ядра, сохраняя свои оболочки, вступают в профазу (См. Профаза), в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности (См. Плоидность); у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие Д. о. — Ксении — проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в Д. о., спермий остальных — дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Лит.: Навашин С. Г., Избр. труды, т. 1, М.— Л., 1951; Магешвар и П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Поддубная Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Steffen К., Fertilisation, в кн.: Maheshwari P. (ed.). Recent advances in the embryology of angiosperms, Delhi, 1963.
Двойное оплодотворение; 1 — у рябчика: один из спермиев (а) в контакте с ядром яйцеклетки, второй (б) — с одним из полярных ядер (второе полярное ядро не изображено); 2 — у подсолнечника: а — пыльцевая трубка; б — синергиды (одна из них повреждена пыльцевой трубкой); в — яйцеклетка; г — спермий в контакте с ядром яйцеклетки; д — центральная клетка; е — второй спермий в контакте со вторичным ядром зародышевого мешка.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .

Опыление и оплодотворение растений. Половое размножение
Половое размножение характерно для большинства растений, за исключением некоторых водорослей.
Опыление — это перенос пыльцы с тычинок на рыльце пестика. Различают перекрестное опыление и самоопыление (рис.1).

Рис.1 Виды опыления цветковых растений
При самоопылении происходит перенос пыльцы с тычинок на рыльце пестика в пределах одного и того же цветка.
В природе самоопыление происходит редко, зачастую еще в бутонах, до раскрытия цветка (пшеница, горох, фасоль, фиалка, томат). Главное преимущество самоопыления — оно не зависит от погодных условий и насекомых, поэтому осуществляется при любых условиях. Не все обоеполые цветки являются самоопыляющимися. Большинство растений дают полноценные семена только при перекрестном опылении.
Опыление, при котором пыльца тычинок одного цветка попадает на рыльце пестика другого, называют перекрестным. Перекрестное опыление осуществляется насекомыми и ветром. Реже — птицами, летучими мышами и водой.
Строение цветков насекомоопыляемых растений разнообразно (вишня, слива, яблоня, сирень, роза и многие другие). Они имеют ярко окрашенный или белый венчик и сильный запах. Цветки крупные или собраны в соцветия.
Запах цветков и их яркая окраска привлекают насекомых. Пчелы, шмели, мухи, бабочки, жуки и муравьи питаются пыльцой и нектаром цветка. Нектарники, расположенные в глубине цветка, выделяют нектар до тех пор, пока цветок не завянет. Тело насекомого, пытающегося добраться до нектарников, обильно покрывается пыльцой. Перелетая с одного цветка на другой в поисках пищи, они переносят прилипшую к их телу пыльцу с тычинок одних цветков на рыльца пестиков других.
Опыление ветром возникло в процессе эволюции как приспособление к неблагоприятным условиям. Надежда на немногочисленных тогда мух, бабочек, пчел и других насекомых была слабой. Позже насекомых стало больше. Но наряду с насекомоопыляемыми растениями, существуют опыляемые ветром. Это многие злаковые травы лугов, степей и саванн, обитатели леса (береза, ольха, осина, дуб, орешник), пустынь и полупустынь (полыни, солянки) (рис.2).
У ветроопыляемых растений бывает очень много пыльцы. Она легкая, сухая и мелкая. Околоцветник отсутствует или плохо развит и не препятствует движению ветра. Перистые рыльца пестиков приспособлены к улавливанию пыльцы. Тычиночные нити длинные и свисающие. Раскачиваясь на ветру, они распыляют зрелую пыльцу.
Большинство ветроопыляемых растений цветет до распускания листьев, что облегчает опыление. Но оно зачастую зависит от погоды. В облачные, дождливые дни осадки смывают пыльцу и тем самым снижают урожай.
Искусственное опыление осуществляет человек, перенося пыльцу с тычинок на рыльца пестиков. Такое опыление требует больших затрат времени и трудно осуществимо на больших площадях. Чаще всего к нему прибегают при выведении новых сортов.
Оплодотворение (рис.2) происходит после опыления.Пыльца, или пыльцевое зерно, попав на рыльце пестика, выделяющего липкую жидкость, прорастает, образуя длинную, тонкую пыльцевую трубку. Пыльцевая трубка, постепенно удлиняясь, продвигается по столбику рыльца по направлению к завязи (нижняя, важнейшая часть пестика). В завязи образуются семязачатки (семяпочки). Снаружи они защищены покровами, а внутри находится зародышевый мешок, состоящий из нескольких клеток.
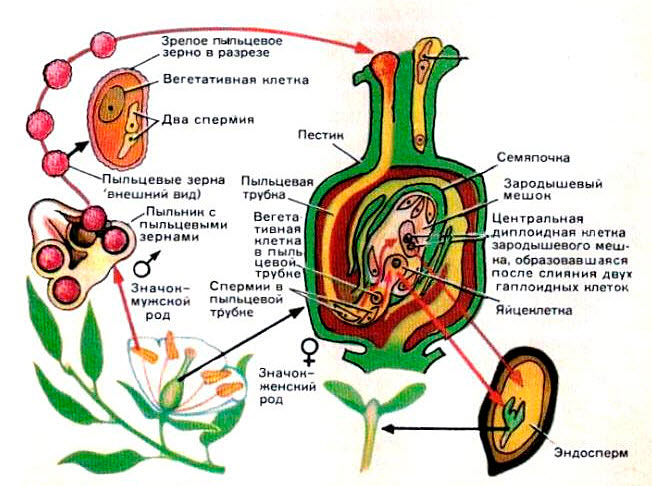
Рис.2 Процесс двойного оплодотворения
Одна из клеток в зародышевом мешке — яйцеклетка, это женская половая клетка (женская гамета). Другая — крупная центральная клетка.
В пыльцевой трубке находятся две маленькие мужские половые клетки (мужские гаметы) — спермии. Когда пыльцевая трубка достигает семязачатка и врастает в него, один спермий сливается с яйцеклеткой. Слияние двух половых клеток (гамет) называется оплодотворением. Из оплодотворенной яйцеклетки впоследствии разовьется зародыш растения. Второй спермий сливается с крупной центральной клеткой. Из нее образуется ткань эндосперм. В клетках этой ткани накапливаются питательные вещества для развития зародыша. Покровы семязачатка превращаются в семенную кожуру. Таким образом два одинаковых спермия сливаются с двумя разными женскими гаметами. Происходит двойное оплодотворение (открыл у лилейных русский ученый в 1898 г. Г.Навашин). После оплодотворения образуется семя, которое состоит из зародыша, запасающей ткани (эндосперма) и семенной кожуры. Из стенки завязи образуется плод.
Опыление — перенос пыльцы на рыльце пестика. Бывает самоопыление — внутри закрытого цветка (горох, пшеница) и перекрестное: насекомыми — цветки яркие, с запахом (яблоня, сирень, роза); ветром — цветки без околоцветника, распускаются до появления листьев, много мелкой легкой пыльцы (береза, дуб, полынь). Искусственное опыление проводит человек. После опыления происходит оплодотворение — процесс слияние мужских и женских половых клеток — гамет. Двойное оплодотворение — два спермия из пыльцы, достигнув завязи пестика по пыльцевой трубке, оплодотворяют две женские гаметы. При слиянии яйцеклетки и спермия образуется зародыш. При слиянии второго спермия и центральной клетки образуется эндосперм (запас питательных веществ). Из покровов семязачатка образуется кожура семени, а из стенки завязи пестика — плод.

(1 слайд)Этот процесс, характерный для всех покрытосеменных. Значение двойного оплодотворения заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения. Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и, следовательно, развивается гораздо быстрее, чем у многих других растений, например у голосеменных.
Явление открыто русским учёным С. Г. Навашиным в 1898 на 2 видах растений — лилии (Lilium martagon) и рябчике (Fritillaria orientalis)
(2 слайд)Гаметы цветковых растений образуются в главных частях цветка – тычинках и пестиках.
(3 слайд)В пыльниках тычинок формируется пыльца, внутри которой развиваются мужские половые клетки.
(4 слайд)Женские половые клетки – яйцеклетки – образуются в семязачатках завязи пестика.
(5 слайд)В результате опыления пыльца попадает на рыльце пестика и прорастает.
Одна пыльцевая клетка вытягивается и образует пыльцевую трубку. В ней по цитоплазме двигаются два спермия.
(6 слайд)Продвигаясь сквозь ткани рыльца и столбика пестика, пыльцевая трубка достигает завязи и прорастает в семязачаток.
(7 слайд)Там спермий сливается с яйцеклеткой, образуя зиготу – из нее сформируется зародыш нового растения.
Второй спермий сливается с центральной клеткой. Образовавшаяся клетка делится и из нее в дальнейшем сформируется эндосперм – питательная ткань для зародыша.
(8 слайд)В результате двойного оплодотворения из семязачатка сформируется семя.
(9 слайд)Из зиготы образуется зародыш семени, из оболочек семязачатка – семенная кожура.
(10 слайд)Биологический смысл двойного оплодотворения весьма велик:
В отличие от голосеменных , триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов. Увеличение уровня плоидности эндосперма до 3n способствует более быстрому росту ткани по сравнению с диплоидными тканями спорофита
Предварительный просмотр:
Подписи к слайдам:
Двойное оплодотворение – Характерно для всех покрытосеменных Заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения Открыто русским учёным С. Г. Навашиным в 1898 на 2 видах растений — лилии ( Lilium martagon ) и рябчике ( Fritillaria orientalis )
Главные части цветка:
Пыльца формируется в пыльниках В пыльце формируются мужские половые клетки
Женские половые клетки (яйцеклетки) образуются в семязачатках в завязи пестика
Биологический смысл двойного оплодотворения весьма велик: В отличие от голосеменных , триплоидный эндосперм образуется лишь в случае оплодотворения С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов Увеличение уровня плоидности эндосперма до 3n способствует более быстрому росту ткани по сравнению с диплоидными тканями спорофита
Двойное оплодотворение, половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений — лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго — с полярными ядрами или со вторичным ядром зародышевого мешка.
Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки — эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой.
Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой — проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое — ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое — мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы.
При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден.
Следствие двойного оплодотворения — ксении — проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных — дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез, а также опыление.
Читайте также: