
Двойное оплодотворение у цветковых растений идиоадаптация
Добавил пользователь Владимир З. Обновлено: 18.09.2024
Уникальная особенность цветковых растений - двойное оплодотворение.
В завязь покрытосеменных растений проникает два спермия, один из них сливается с яйцеклеткой, дав начало диплоидному зародышу. Другой соединяется с центральной диплоидной клеткой. Образуется триплоидная клетка, из которой возникнет эндосперм - питательный материал для развивающегося зародыша ( рис. 77 ). Этот процесс, характерный для всех покрытосеменных, открыт в конце прошлого века С.Г. Навашиным и получил название двойного оплодотворения. Значение двойного оплодотворения, по- видимому, заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения. Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и, следовательно, развивается гораздо быстрее, чем у многих других растений, например у голосеменных.
У цветковых растений имеется ряд особенностей образования половых клеток и оплодотворения. Оплодотворению у них предшествует образование сильно редуцированного гаплоидного поколения - гаметофитов. После оплодотворения прорастание пыльцы цветковых растений начинается с разбухания зерна и образования пыльцевой трубки , которая прорывает спородерму в более тонком ее месте - так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика , в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия . В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий ( нуцеллус ) через микропиле семязачатка , реже - иным образом. Проникнув в зародышевый мешок , пыльцевая трубка разрывается, и ее содержимое изливается внутрь. Один изспермиев сливается с яйцеклеткой, и образуется диплоидная зигота, дающая затем начало зародышу . Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка , что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Весь этот процесс получил название двойного оплодотворения. Он был впервые описан в 1898 г. выдающимся русским цитологом и эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка - антиподы и синергиды в оплодотворении не участвуют и довольно быстро разрушаются.
Как и у других высших растений , при мейозе у цветковых образуются споры . Микроспоры (мужские споры) образуются в пыльниках , из них формируются пыльцевые зерна (мужские гаметофиты). Зрелое пыльцевое зерно состоит из трех клеток - вегетативной (или клетки, пыльцевой трубки ) и двух спермиев(лишенные жгутика сперматозоиды). Мегаспоры (женские споры) формируются в семяпочках внутри завязи пестика .
Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных , где довольно мощный гаплоидный эндосперм развивается независимо от процесса оплодотворения, у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов. Увеличение же уровня плоидности эндосперма до 3n, по-видимому, способствует более быстрому росту этой полиплоидной ткани по сравнению с диплоидными тканями спорофита .
Взаимодействие пыльцевой трубки гаметофита с тканями спорофита - сложный процесс, регулируемый химическими веществами. Так, выяснилось, что если промыть пыльцу дистиллированной водой, она теряет способность к прорастанию. Если сконцентрировать полученный раствор и обработать концентратом пыльцу, она вновь станет полноценной. После прорастания рост пыльцевой трубки контролируется тканями пестика. Например, у хлопчатника рост трубки до яйцеклетки занимает 12-18 ч, но уже через 6 ч можно установить, к какой семяпочке направляется пыльцевая трубка: в этой семяпочке начинается разрушение особой клетки - синергиды . Как растение направляет рост трубки в нужную сторону и каким образом синергида узнает о ее приближении, пока еще не известно.
Во многих случаях у цветковых растений существует "запрет" на самоопыление: спорофит "узнает" своего мужского гаметофита и не разрешает ему участвовать в оплодотворении. В некоторых случаях при этом собственная пыльца не прорастает на рыльце пестика. В большинстве же случаев рост пыльцевой трубки начинается, но затем останавливается и она не достигает яйцеклетки. Например, у первоцвета весеннегоеще Ч.Дарвин обнаружил две формы цветков - длинностолбиковые (с длинным столбиком и короткими тычинками) и короткостолбиковые (столбик короткий, тычиночные нити длинные). У короткостолбиковых растений пыльца почти вдвое крупнее, а клетки сосочков рыльца мелкие. Все эти признаки контролируются группой тесно сцепленных генов.
Опыление эффективно только при переносе пыльцы с одной формы на другую. За распознавание своей пыльцы отвечают молекулы-рецепторы, представляющие собой сложные комплексы белков с углеводами. Показано, что растения дикой капусты , которые не вырабатывают в тканях рыльца молекул рецептора, могут самоопыляться. У нормальных растений рецепторы появляются на рыльце за день до открытия цветка. Если раскрыть бутон и нанести на него собственную пыльцу за два дня до распускания, то оплодотворение произойдет, а если за один день до распускания - то нет.
Интересно, что в некоторых случаях самонесовместимость пыльцы у растений определяется серией множественных аллелей одного гена, сходно с несовместимостью при пересадках тканей у животных. Эти аллели обозначаются буквой S, и число их в популяции может достигать десятков и даже сотен. Если, например, генотип производящего яйцеклетки растения - s1s2, а производящего пыльцу - s2s3, то прорастать при перекрестном опылении будут только 50% пылинок - те, что несут аллель s3. При наличии десятков аллелей большая часть пыльцы при перекрестном опылении нормально прорастает, а самоопыление полностью предотвращается.
Перенос пыльцы из пыльника на рыльце пестика называется опылением. Различают два вида опыления: перекрестное и самоопыление.
При самоопылении рыльце принимает пыльцу того же цветка либо другого, но той же особи. Возможно опыление в закрытых, нераспустившихся цветках (горох). При перекрестном опылении переносится пыльца от разных особей. Это основной тип опыления цветковых растений (яблоня, ива, огурец и др.).

Перекрестное опыление
Перекрестное опыление осуществляется естественным (насекомыми, птицами, летучими мышами, ветром, водой) и искусственным (производит человек) путями.
Приспособленность растений к опылению ветром проявляется в наличии голых цветков, либо невзрачных, слабо развитых околоцветников. Они лишены нектарников и запаха, пыльцы образуют много, она легкая, сухая, мелкая, рыльца длинные, с большой поверхностью для улавливания пыльцы (рожь, кукуруза).
Приспособленность растений к опылению насекомыми характеризуется яркой окраской венчика, наличием нектарников, запаха (одуванчик, земляника). Пищей для насекомых являются нектар и пыльца. Окраска и запах служат для привлечения опылителей. Иногда цветки обладают запахом, характерным для самок насекомых того же вида. Это привлекает к ним самцов, которые и осуществляют опыление. Эволюция цветковых растений и их опылителей шла параллельно. Это так называемая сопряженная эволюция.

Приспособление растений к опылению насекомыми и ветром
Перекрестное опыление обеспечивает обмен генами, поддерживает высокую гетерозиготность популяций, дает материал для естественного отбора и сохраняет самое выносливое потомство — носителей наиболее благоприятного сочетания генов.
Искусственное опыление
Искусственное опыление производит человек для повышения урожая или получения новых сортов растений. При этом для нанесения пыльцы на рыльце пестика используют разные способы. Так, у кукурузы, имеющей однополые цветы, пыльцу собирают, стряхивая верхушечные метелки мужских цветков в бумажные воронки. Затем собранной пыльцой посыпают выступающие на верхушке початка длинные рыльца женских цветков.
При искусственном опылении подсолнечника стебли двух соседних растений наклоняют так, чтобы можно было прижать цветущую поверхность одной корзинки к другой. Можно переносить пыльцу, поочередно прижимая руку в варежке из мягкой материи к цветущим корзинкам разных растений.

Схема искусственного опыления
Для получения новых сортов растений с обоеполыми цветками необходима подготовка к искусственному опылению. Прежде всего из цветков растения, избранного в качестве материнского, еще в бутоне удаляют пыльники и защищают эти цветки марлевыми или бумажными мешочками от попадания пыльцы. Через 2-3 дня, когда бутоны раскроются, наносят на рыльца пестиков заготовленную пыльцу другого сорта чистой сухой акварельной кисточкой, мягким поролоном или кусочком резинки, прикрепленными к проволоке.
Двойное оплодотворение у цветковых растений
После опыления происходит процесс оплодотворения, но для этого нужен ряд условий: пыльца должна не только удержаться на рыльце, но и прорасти через столбик, достигнуть семязачатка и обеспечить слияние мужских клеток с женскими.
Двойное оплодотворение характерно для цветковых растений.
Обычно на рыльце попадает множество пыльцевых зерен. Они, как правило, имеют шероховатую поверхность и удерживаются липкой кожицей рыльца. Кроме этого, при попадании совместимой пыльцы клетки рыльца выделяют вещества, стимулирующие ее прорастание.

Схема двойного оплодотворения у цветковых растений
Начинается прорастание пыльцевых зерен с набухания. Затем через специальные поры (каналы) в наружной оболочке пыльцевого зерна внутренняя выпячивается в тонкую пыльцевую трубку, куда переходят вегетативное ядро и спермин. Пыльцевые трубки всех совместимых зерен, удержавшихся на рыльце пестика, растут по столбику, направляясь к семязачатку. Одна из них обгоняет в росте другие и, достигнув пыльцевхода, проникает через него к зародышевому мешку и здесь изливает в него свое содержимое.
Один из спермиев сливается с яйцеклеткой, а другой — со вторичным ядром центральной диплоидной клетки. Вегетативное ядро разрушается еще до проникновения пыльцевой трубки в зародышевый мешок.
Двойное оплодотворение у цветковых растений открыл русский цитолог и эмбриолог растений С.Г.Навашиным в 1898г.
При наличии в завязи семязачатков в каждом из них происходит вышеописанный процесс двойного оплодотворения. Называется он двойным потому, что сливаются две мужские клетки с двумя клетками женского гаметофита. В дальнейшем после оплодотворения в цветке начинается развитие семени и плода.
Образование семян
После оплодотворения внутри зародышевого мешка начинается быстрое митотическое деление триплоидного вторичного ядра, не имеющего периода покоя. Образуется большое количество ядер, затем между ними возникают, перегородки.
Эти вновь образовавшиеся клетки продолжают деление, заполняя всю полость зародышевого мешка питательной тканью — эндоспермом, который у одних растений полностью расходуется во время развития зародыша (бобовые, тыквенные), а у других — сохраняется в зрелых семенах (злаки). Одновременно происходит разрастание зародышевого мешка и семяпочки.
Формирование зародыша начинается с деления зиготы. После периода покоя зигота делится митотически на две клетки. Верхняя клетка, прилегающая к пыльцевходу, образует подвесок, отодвигающий нижнюю клетку в глубь эндосперма. Подвесок у одних видов растений остается одноклеточным, у других — делится поперечными перегородками и становится многоклеточным. Нижняя клетка разрастается в предзародыш семени сферической формы. Предзародыш делится на 4 клетки двумя перпендикулярными перегородками, затем каждая из этих клеток делится еще на две.
Сначала клетки более или менее однородны. По мере дальнейшего деления происходит дифференцировка клеток на зачаточный корешок, зачаточный стебель, зачаточные листочки (семядоли) и зачаточную почечку, окруженную семядолями. К этому времени семяпочка превращается в семя, ее покровы и остатки эндосперма образуют кожицу семени.
Таким образом, из оплодотворенной диплоидной яйцеклетки формируется зародыш семени, а из вторичной триплоидной клетки — питательная ткань — эндосперм, покровы семязачатка превращаются в покровы семени, а стенка завязи, разрастаясь, образует околоплодник.

Оплодотворение – это процесс слияния двух клеток, в результате чего происходит образование новой клетки, дающей начало|начало другому организму этого же рода|рода или вида. Что такое двойное оплодотворение у цветковых растений и как оно происходит, читайте в данной статье.
Сущность оплодотворения
Оно происходит в результате слияния двух клеток, женской и мужской, и возникновения диплоидной зиготы. В каждой паре хромосом присутствует одна отцовская и одна материнская клетка. Сущность процесса оплодотворения заключается в том, чтобы восстановить диплоидный набор хромосом и объединить наследственный материал родителей. Их потомство будет более жизнеспособным, так как соединит в себе самые полезные качества от отца и матери.
Оплодотворение — что такое?
Это процесс побуждения яйца|яйца к развитию в результате объединения ядер. Оплодотворение — что такое? Это необратимый процесс, который происходит в результате слияния разнополых гамет и объединения их ядер. Оплодотворённое яйцо не подвергается этой процедуре второй раз.
Но существуют растения, которые воспроизводят новое поколение только при помощи женской гаметы без оплодотворения. Такое размножение называется девственным. Примечательно, что эти два способа размножения у одного вида растений могут чередоваться.
Двойное оплодотворение цветковых растений
Половые клетки обоих начал называются гаметами. Причём женскими являются яйцеклетки, а мужскими – спермии, которые у растений семенных неподвижные, а у споровых – подвижные|подвижные. Оплодотворение — что такое? Это появление особой клетки – зиготы, содержащей наследственные признаки спермия и яйцеклетки.
Цветковые растения обладают сложным оплодотворением, которое называется двойным, поскольку, кроме яйцеклетки, оплодотворяется ещё одна особая клетка. Формирование спермий происходит в пылинках пыльцы, а их созревание осуществляется в тычинках, точнее в их пыльниках. Местом образования яйцеклеток являются семязачатки, расположенные в завязи|завязи пестика. Когда яйцеклетка оплодотворится спермием, из семязачатка начинают развиваться семена|семёна.
Чтобы оплодотворение у цветковых произошло, сначала нужно опылить растение, то есть на рыльце пестика должны попасть пылинки пыльцы. Оказавшись на рыльце, они начинают прорастать внутрь завязи|завязи, в результате чего образуется пыльцевая трубка. Одновременно с этим в пылинке происходит образование двух спермиев. Они не стоят|стоят на месте, а начинают продвигаться к пыльцевой трубке, которая проникает в семязачаток. Здесь в результате деления и удлинения одной клетки происходит образование зародышевого мешка.
Он нужен для расположения в нём яйцеклетки и ещё одной клетки, в которой сосредоточен двойной набор наследственной информации. После этого происходит прорастание пыльцевой трубки в зародышевый мешок и слияние одного спермия с яйцеклеткой, в результате которого образуется зигота, а другого – с клеткой особой. Развитие зародыша происходит из зиготы. Второе слияние образует питательную ткань, или эндосперм, необходимый для питания зародыша в период роста|роста.
Что нужно для существования каждого вида растений?
- Прежде всего необходимо восстановить диплоидный набор хромосом, а в его пределах — их парность.
- Обеспечить материальную непрерывность между поколениями, следующими чередой.
- Объединить в одном виде или роде наследственные свойства двух родителей.
Всё|Все это осуществляется на генетическом уровне. Для того чтобы оплодотворение осуществилось, созревание материнских и отцовских гамет должно произойти одновременно.
Оплодотворение у покрытосеменных растений
Этот процесс впервые охарактеризовал немецкий учёный Страсбургер во второй половине девятнадцатого века|века. Оплодотворение покрытосеменных растений происходит в результате слияния двух ядер разных гамет: с мужским и женским началом. Их цитоплазма не участвует в оплодотворении. Собственно оплодотворение происходит тогда, когда спермий сливается с ядром яйцеклетки.
Местом возникновения спермиев является пыльцевое зерно или пыльцевая труба. Зерно начинает прорастать после того, как попадает|попадает на рыльце. Время начала|начала этого процесса у каждого растения разное, как и время оплодотворения. Например, пыльцевые зерна|зёрна свёклы прорастают через два часа, а кукурузы – моментально. Первый признак прорастания зерна|зёрна – его увеличение в объёме. Обычно одно пыльцевое зерно образует одну трубку. Но некоторые растения не подчиняются этому правилу и образуют несколько трубок, из которых только одна достигает своего развития.
Пыльцевая трубка с передвигающимися по ней спермиями растёт и в конце концов разрывается. Всё|Все её содержимое оказывается внутри зародышевого мешка. Один из проникших сюда спермиев внедряется в яйцеклетку и сливается с её гаплоидным ядром. Оплодотворение — что такое? Это слияние двух ядер: спермия и яйцеклетки. Оплодотворённая яйцеклетка начинает делиться, получаются две новые клетки. Они делятся на четыре и так далее. Таким образом, происходит многократное деление, в результате которого развивается зародыш растения.
Покрытосеменные растения после процесса оплодотворения обладают способностью развивать дополнительный орган|орган, который называется эндоспермом. Это не что иное, как питательная среда зародыша. При слиянии второго спермия и диплоидного ядра|ядра происходит образование определённого набора хромосом, из которых два – материнского происхождения, и один – отцовского. Таким образом, двойное оплодотворение организмов растительного происхождения осуществляется тогда, когда один спермий сливается с яйцеклеткой, а другой – с ядром клетки, расположенной в центре.
Отличительные черты покрытосеменных растений
Благодаря перечисленным признакам покрытосеменные растения занимают господствующее положение в мире.
Особенность оплодотворения покрытосеменных растений
Она вытекает из того, что эти растения имеют двойное оплодотворение. Уникальная особенность представлена явлением, называемым ксениями. Его смысл заключается в том, что пыльца напрямую влияет на свойства и признаки эндосперма. Для примера возьмём кукурузу.
Она бывает с жёлтыми и белыми семенами|семёнами. Их цвет зависит от оттенка эндосперма. При опылении женских цветков белозёрной кукурузы пыльцой желтозёрного сорта|сорта её окраска всё равно будет жёлтой, хотя развитие эндосперма происходит на растении с белыми зёрнами.
Какую роль играют цветковые растения?
Эти растения насчитывают 13 000 родов|родов и 250 000 видов. Они получили широкое распространение по всему миру. Цветковые растения – это ключевые компоненты биосферы, производящие органические вещества, связывающие углекислоту и выделяющие кислород. Пастбищные цепи питания начинаются именно с них. Многие разновидности цветковых растений человек использует в пищу|пищу. Из них строят жилища|жилища и изготавливают различные хозяйственные материалы.
Не обходится без них и медицина. Отдельные виды покрытосеменных растений являются господствующими на планете, им отводится решающая роль в формировании покрова|покрова растительности и создании основной части наземной фитомассы. В конечном итоге именно этими растениями определяется возможность самого|самого существования человека на земле как вида биологического.
Процесс оплодотворения у цветковых растений

Пояснение.
Процесс оплодотворения у цветковых растений характеризуется слиянием спермия с центральной клеткой и слиянием спермия и яйцеклетки; образованием зиготы в зародышевом мешке
В семязачатке к этому времени бывает развит зародышевый мешок, образовавшийся из мегаспоры и состоящий из семи клеток, одна из которых — яйцеклетка (женская гамета), а самая крупная — центральная клетка с двумя ядрами.
Когда пыльцевая трубка дорастает до зародышевого мешка и входит в него, она лопается и спермин осуществляют оплодотворение — один сливается с яйцеклеткой, образуя зиготу, а другой — с центральной клеткой. Этот процесс называют двойным оплодотворением. Из зиготы затем развивается зародыш, а из центральной клетки эндосперм (ткань, запасающая питательные вещества). После оплодотворения из семязачатков формируются семена|семёна, а сам цветок превращается в плод.
Видео по теме : Процесс оплодотворения у цветковых растений
Двойное оплодотворение у растений имеет большое биологическое значение. Оно было открыто Навашиным в 1898 г. Далее рассмотрим подробнее, как происходит двойное оплодотворение у растений.
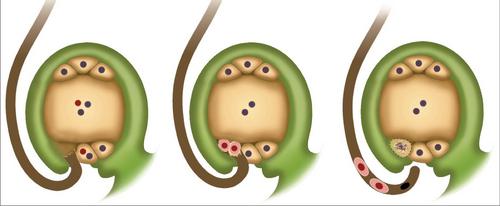
Биологическое значение
Процесс двойного оплодотворения способствует активному развитию питательной ткани. В связи с этим семяпочка не запасает вещества впрок. Это, в свою очередь, объясняет ее быстрое развитие.
Схема двойного оплодотворения
Коротко явление можно описать следующим образом. Двойное оплодотворение у покрытосеменных растений состоит в проникновении в завязь двух спермиев. Один сливается с яйцеклеткой. Это способствует началу развития диплоидного зародыша. Второй спермий соединяется с центральной клеткой. В результате формируется триплоидный элемент. Из этой клетки появляется эндосперм. Он является питательным материалом для развивающегося зародыша.
Развитие пыльцевой трубки
Двойное оплодотворение у покрытосеменных начинается после образования гаплоидного сильно редуцированного поколения. Оно представлено гаметофитами. Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.

Формирование клеток: общие сведения
Процесс двойного оплодотворения цветковых растений осуществляется особыми половыми клетками. Их формирование происходит в два этапа. Первая стадия называется спорогенез, вторая – гематогенез. В случае образования мужских клеток эти этапы именуются микроспорогенез и микрогематогенез. При образовании женских половых элементов приставка меняется на "мега" (или "макро"). Спорогенез основывается на мейозе. Это процесс формирования гаплоидных элементов. Мейозу, так же как и у представителей фауны, предшествует размножение клеток посредством митотических делений.
Образование спермиев
Первичное формирование мужских половых элементов осуществляется в особой ткани пыльника. Она называется археспориальной. В ней в результате митозов происходит формирование многочисленных эелементов – материнских клеток пыльцы. Они и вступают затем в мейоз. Вследствие двух мейотических делений образуется 4 гаплоидные микроспоры. Некоторое время они лежат рядом, формируя тетрады. После этого происходит их распад на пыльцевые зерна - отдельные микроспоры. Каждый из образованных элементов начинает покрываться двумя оболочками: внешней (экзина) и внутренней (интина). Затем начинается следующий этап – микрогаметогенез. Он, в свою очередь, состоит из двух митотических последовательных делений. После первого формируется две клетки: генеративная и вегетативная. Впоследствии первая проходит еще одно деление. В результате образуется две мужские клетки – спермии.

Макроспорогенез и мегаспорогенез
В тканях семяпочки начинает обособляться один или несколько археспориальных элементов. Они начинают усиленно расти. Вследствие такой активности они становятся значительно крупнее остальных клеток, окружающих их в семяпочке. Каждый археспориальный элемент один, два или более раз подвергается делению митозом. В некоторых случаях клетка может сразу трансформироваться в материнскую. Внутри нее происходит мейоз. В результате него формируется 4 гаплоидные клетки. Как правило, самая крупная из них начинает развиваться, превращаясь в зародышевый мешок. Три оставшиеся постепенно дегенерируют. На данном этапе макроспорогенез завершается, начинается макрогематогенез. В ходе него происходят митотические деления (у большей части покрытосеменных их три). Цитокинез не сопровождает митозы. В результате трех делений формируется зародышевый мешок с восемью ядрами. Они впоследствии обосабливаются в самостоятельные клетки. Эти элементы распределяются определенным образом по зародышевому мешку. Одна из обособленных клеток, которая, собственно, является яйцеклеткой, совместно с двумя другими – синергидами, занимает место у микропиле, в которое осуществляется проникновение спермиев. В этом процессе синергиды исполняют очень значимую роль. В них содержатся ферменты, которые способствуют растворению оболочек на пыльцевых трубках. В противоположной стороне зародышевого мешка располагаются другие три клетки. Они именуются антиподами. С помощью этих элементов происходит передача из семяпочки питательных веществ в зародышевый мешок. Оставшиеся две клетки располагаются в центральной части. Зачастую они сливаются. В результате их соединения формируется диплоидная центральная клетка. После того как произойдет двойное оплодотворение, и в завязь проникнут спермии, один из них, как выше сказано было, сольется с яйцеклеткой.

Особенности пыльцевой трубки
"Запрет" на самоопыление
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. "Запрет" на самоопыление проявляется в том, что спорофит "идентифицирует" собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же – короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца – мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.
Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев "самонесовместимость" пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, - s1s2, а вырабатывающего пыльцу - s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.

В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
Читайте также: