
Чем представлен гаметофит высших споровых растений
Добавил пользователь Дмитрий К. Обновлено: 18.09.2024
Включают следующие ныне существующие отделы: моховидные (Bryophyta), плауновидные (Lycopodiophyta), псилотовидные (Psilotophyta), хвощевидные (Equisetophyta), папоротниковидные (Polypodiophyta).
Споровые растения появились в конце силурийского периода, более 400 млн. лет тому назад. Первые представители споровых были небольших размеров и имели простое строение, но уже у примитивных растений наблюдалась дифференциация на элементарные органы. Усовершенствованию органов соответствовало усложнение внутреннего строения и онтогенеза. В жизненном цикле происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено диплоидным спорофитом, половое – гаплоидным гаметофитом.
На спорофитеобразуются спорангии, внутри которых в результате мейотического деления образуются гаплоидные споры. Это мелкие, одноклеточные образования, лишенные жгутиков. Растения, у которых все споры одинаковые, называются равноспоровыми. У более высокоорганизованных групп споры двух типов: микроспоры (формируются в микроспорангиях), мегаспоры (формируются в мегаспорангиях). Это разноспоровые растения. При прорастании споры образуется гаметофит.
Полный жизненный цикл (от зиготы до зиготы) состоит из гаметофита(период от споры до зиготы) и спорофита (период от зиготы до образования спор). У плаунов, хвощей и папоротников эти фазы представляют собой как бы отдельные физиологически самостоятельные организмы. У мхов гаметофит является самостоятельной фазой жизненного цикла, а спорофит сведен до его своеобразного органа – спорогона(спорофит живет на гаметофите).
На гаметофите развиваются органы полового размножения: архегонии и антеридии. В архегониях, похожих на колбу, образуются яйцеклетки, а в мешковидных антеридиях - сперматозоиды. У равноспоровых растений гаметофиты обоеполые, у разноспоровых – однополые. Оплодотворение происходит лишь при наличии воды. При слиянии гамет образуется новая клетка – зигота с двойным набором хромосом (2n).
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса: Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida)
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha) (рис 11. 1).
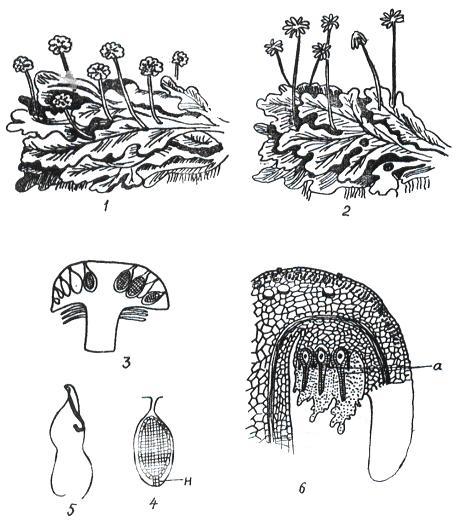
Рис. 11. 1. Цикл воспроизведения маршанции: 1– таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6– вертикальный разрез через женскую подставку (а – архегоний).
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии. На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).
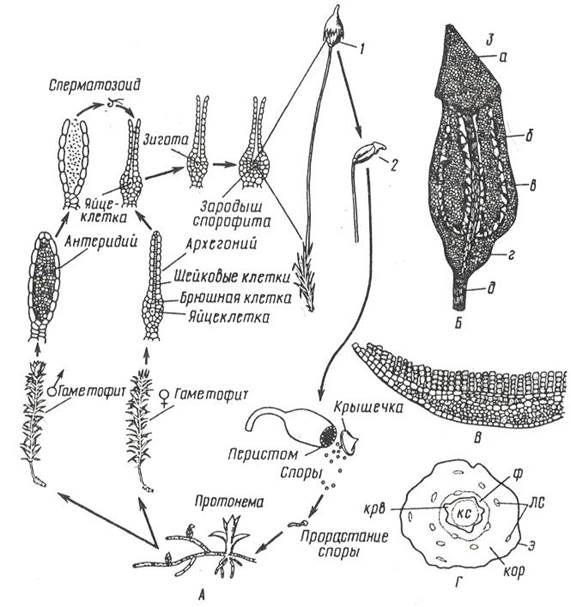
Рис. 11. 2. Кукушкин лен: А– цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум(Sphagnum) (рис. 11. 3).
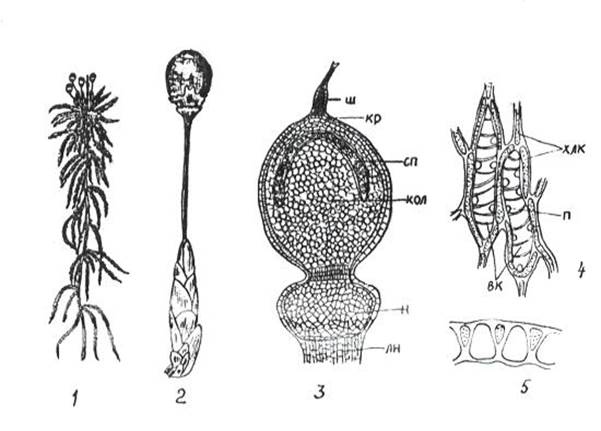
Рис 11. 3. Сфагнум: 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон ( ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа ( хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы. У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун(Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный (L. clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum) (рис. 11. 4).

Рис. 11. 4. Плаун булавовидный.
Родбаранец(Huperzia).Представитель рода -баранец обыкновенный (H. selago) распространен в тундровой, лесотундровой и северной лесной зонах и произрастает в южно-таежных еловых лесах и ольшаниках, а также в мшистых лесах и на альпийских лугах.
Роддифазиаструм (Diphasiastrum). Представитель рода дифазиаструм сплюснутый (D. complanatum) растет на сухих песчаных почвах в сосновых лесах.
Цикл развития на примере плауна булавовидного (рис. 11. 5).
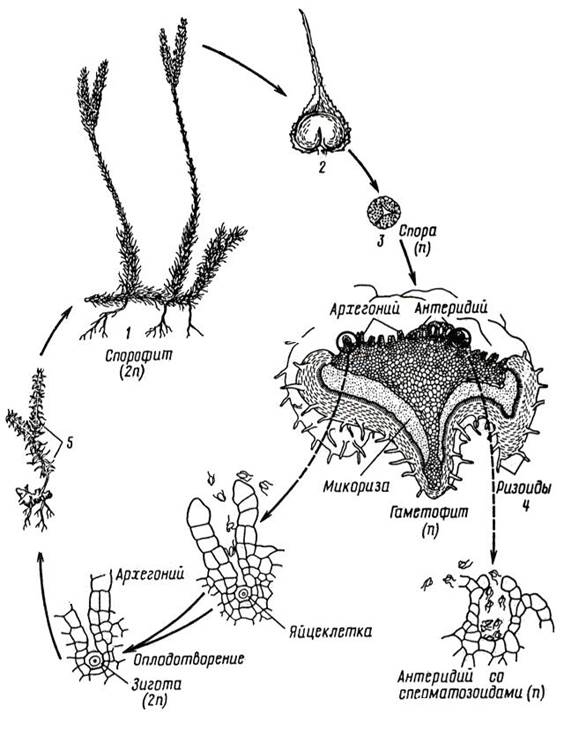
Рис. 11. 5. Цикл развития плауна булавовидного:1 – спорофит; 2 – спорофилл со спорангием; 3 – спора; 4 – гаметофит с антеридиями и архегониями; 5 – развивающийся на гаметофите из зародыша молодой спорофит.
Стелющиеся побеги плауна булавовидного достигают до 25 см в высоту и более 3 м в длину. Стебли покрыты спирально расположенными ланцетно-линейными мелкими листьями. В конце лета на боковых побегах образуется обычно два спороносных колоска. Каждый колосок состоит из оси и мелких тонких спорофиллов – видоизмененных листьев, в основании которых расположены спорангии почковидной формы.
В спорангиях после редукционного деления клеток спорогенной ткани образуются одинаковой величины, одетые толстой желтой оболочкой гаплоидные споры. Они прорастают после периода покоя через 3-8 лет в обоеполые заростки, представляющие собой половое поколение и живущие сапротрофно в почве, в виде клубенька. От нижней поверхности отходят ризоиды. Через них в заросток врастают гифы гриба, образуя микоризу. В симбиозе с грибом, который обеспечивает питание, живет заросток, лишенный хлорофилла и неспособный к фотосинтезу. Заростки многолетние, развиваются очень медленно, лишь через 6-15 лет на них образуются архегонии и антеридии. Оплодотворение совершается при наличии воды. После оплодотворения яйцеклетки двужгутиковым сперматозоидом образуется зигота, которая без периода покоя прорастает в зародыш, развивающийся во взрослое растение.
В официнальной медицине споры плауновприменялись в качестве детской присыпки и обсыпки для пилюль. Побеги баранца обыкновенного используют для лечения больных, страдающих хроническим алкоголизмом.
Класс Полушниковые (Isoetopsida)
Селагинелла (Selaginella) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыре мегаспорыв мегаспорангиях и многочисленные микроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.
Отдел Хвощевидные – Equisetophyta
Появились хвощи в верхнем девоне, достигли наибольшего разнообразия в карбоне, когда древесный ярус заболоченных тропических лесов в значительной степени состоял из древовидных хвощевидных, вымерших к началу мезозоя. Современные хвощевидные появились на Земле с мелового периода.
До настоящего времени сохранился лишь один род – хвощ (Equisetum), представленный 30-35 видами, распространенными на всех континентах.
У всех видов хвоща стебли имеют членистое строение с выраженным чередованием узлов и междоузлий. Листья редуцированы до чешуй и располагаются мутовками в узлах. Здесь же образуются и боковые ветви. Ассимилирующую функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток эпидермы пропитаны кремнеземом. Подземная часть представлена сильно развитым корневищем, в узлах которого формируются придаточные корни. У хвоща полевого(Equisetum arvense) боковые ветви корневища служат местом отложения запасных веществ, а также органами вегетативного размножения (рис. 11. 6).

Рис. 11. 6. Хвощ полевой: а, б – вегетативный и спороносный побеги спорофита; в – спорангиофор со спорангиями; г, д – споры; е – мужской гаметофит с антеридиями; ж – сперматозоид; з – обоеполый гаметофит; и - архегонии.
Весной на обычных или специальных спороносных стеблях образуются колоски, состоящие из оси, которая несет особые структуры, имеющие вид шестигранных щитков (спорангиофоры). Последние несут 6-8 спорангиев. Внутри спорангиев образуются споры, одетые толстой оболочкой, снабженной гигроскопичными лентовидными выростами – элатерами. Благодаря элатерамспоры сцепляются вместе в комки, хлопья. Групповое распространение спор способствует тому, что при их прорастании разнополые заростки оказываются рядом, и это облегчает оплодотворение.
Заростки имеют вид небольшой длинно-лопастной зеленой пластинки с ризоидами на нижней поверхности. Мужские заростки меньше женских и несут по краям лопастей антеридии с многожгутиковыми сперматозоидами. На женских заростках в средней части развиваются архегонии. Оплодотворение происходит при наличии воды. Из зиготы развивается зародыш нового растения – спорофита.
В настоящее время большой роли в образовании растительного покрова хвощи не имеют. В лесах, на избыточно увлажненной почве, широко распространен хвощ лесной (E. sylvaticum) с сильно ветвящимися, поникающими боковыми веточками. На лугах, паровых полях, в посевах встречается трудно искореняемый сорняк хвощ полевой (E. arvense). У этого хвоща рано весной появляются неветвящиеся побеги, несущие спороносные колоски. Позднее от корневища развиваются зеленые вегетативные побеги. В лесной зоне на песчаных почвах и в оврагах широко распространен хвощ зимующий(E. hyemale).
Вегетативные побеги хвоща полевого (E. arvense) в официнальной медицине применяют: в качестве мочегонного средства при отеках на почве сердечной недостаточности; при заболеваниях мочевого пузыря и мочевыводящих путей; как кровоостанавливающее средство при маточных кровотечениях; при некоторых формах туберкулеза.
Отдел Папоротниковидные – Polypodiophyta
Возникли папоротники в девоне, когда древовидные папоротники вместе с ныне ископаемыми плаунами и хвощами господствовали в растительном покрове земли. Большинство их вымерло, оставшиеся дали начало мезозойским формам, которые были представлены очень широко. Папоротники намного превосходят по числу современных видов все остальные отделы высших споровых (около 25000).
В лесах Австралии, Южной Америки, Азии произрастают древовидные представители с колонновидными, неветвящимися, до 20 метров высоты стволами. В средней полосе нашей страны папоротники – это многолетние корневищные травы. Многие папоротники, как и мхи, являются индикаторами почв и типов леса. В светлых лесах, на песчаных или сухих подзолистых почвах распространен орляк обыкновенный(Pteridium aquilinum); на влажных богатых почвах кочедыжники(Athyrium) и крупныелесныещитовники (Dryopteris)(рис. 11. 7).
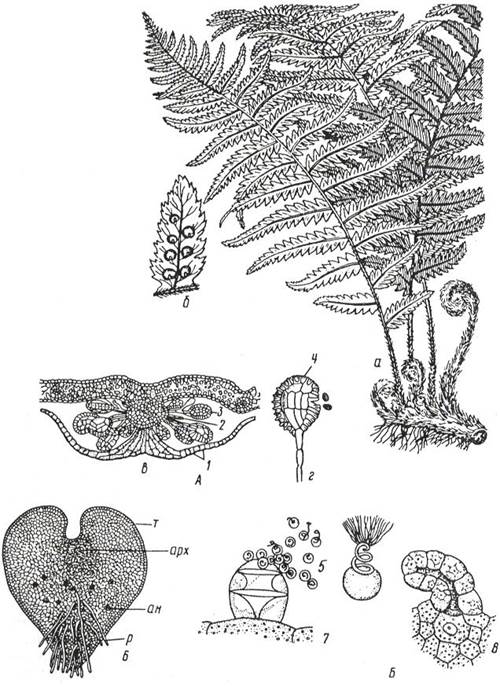
Рис. 11. 7. Щитовник мужской: А – спорофит: а – общий вид; б – сорусы на нижней стороне вайи; в – разрез соруса (1 – индузий, 2 – планцента, 3 – спорангий); г – спорангий (4 – кольцо); Б– гаметофит: 5 – сперматозоиды; 6 – заросток с нижней стороны (т – таллом, р – ризоиды, арх - архегонии, ан – антеридии); 7 – выход сперматозоидов из антеридия; 8 – архегоний с яйцеклеткой.
Цикл развития равноспоровых папоротников
В середине лета на нижней стороне зеленых листьев (у некоторых на специальных спороносных листьях) в виде коричневых бородавочек появляются группы спорангиев (сорусы). Сорусы у многих папоротников прикрыты сверху своеобразным покрывалом - индузием.Спорангии образуются на особом выросте листа (плаценте)и имеют чечевицеобразную форму, длинные ножки и многоклеточные стенки. В спорангиях хорошо выражено механическое кольцо, которое имеет вид узкой несмыкающейся полосы, опоясывающей спорангий. Когда кольцо подсыхает, стенки спорангия разрываются, и споры высыпаются.
Споры, образовавшиеся в спорангиях, одноклеточны и имеют толстую оболочку. При созревании они разносятся током воздуха и в благоприятных условиях прорастают, образуя сердцевидную зеленую многоклеточную пластинку (заросток), прикрепляющуюся к почве ризоидами. Заросток представляет собой половое поколение папоротников (гаметофит). На нижней стороне заростка образуются антеридии (со сперматозоидами) и архегонии (с яйцеклетками). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетки. Из зиготы развивается зародыш, имеющий все основные органы (корень, стебель, лист и особый орган – ножку, прикрепляющую его к заростку). Постепенно зародыш начинает существовать самостоятельно, а заросток отмирает.
У разноспоровых папоротников гаметофиты редуцированы до микроскопических размеров (особенно мужские).
Из корневищпапоротника мужского(Dryopteris filix-mas), получают густой экстракт, который является эффективным противоглистным средством (ленточные черви).
Это растения, которые первыми освоили сушу. Их относят в высшим, т.к. по сравнению с водорослями, у них уже появились ткани и органы - приспособления для жизни в наземной среде.
Водоросли тоже размножаются спорами, но это водные организмы, самые простые по организации.
Основная особенность споровых - размножение специальными клетками - спорами.
Прежде, чем мы разберем, что из себя представляют споры, давайте вспомним, какие бывают клетки.
Клетки:
-
соматические (или вегетативные) - клетки тела организма. У них двойной (диплоидный) набор хромосом, обозначается как 2n
-
половые ( или генеративные, или гаметы) - это “половинчатые” клетки - их набор хромосом в 2 раза меньше, чем у соматических - 1n

Получается, что споры - это такие половые клетки ( или группы клеток) у растений. Растение, которое их образует - спорофит - оно диплоидное (2n)
У животных гаметы образуются путем мейоза, у растений - путем митоза.
У животных гаметы тут же сливаются, происходит оплодотворение, новый организм получает двойной - диплоидный (2n) набор хромосом.
У споровых растений клетка споры прорастает в целый самостоятельный организм - гаметофит, который может опять производить половые клетки - гаметы.

Гаметы сливаются и образуют опять спорофит.
Получается, что жизнь споровых растений - своеобразный цикл - чередование спорофита и гаметофита.
Спорофит — диплоидная многоклеточная фаза в жизненном цикле растений и водорослей, развивающаяся из оплодотворённой яйцеклетки или зиготы и производящая споры.
Архего́ний, или Архегония (др.-греч. ἀρχή — начало, γονή — потомство) — женский орган полового размножения у высших споровых (мохообразные, папоротниковидные и пр.) и голосеменных растений, многоклеточный орган полового размножения гаметофитов; гаметангий, в котором развиваются женские гаметы — яйцеклетки.
Гаметангий — орган полового размножения у растений, в котором развиваются половые клетки. У водорослей и грибов гаметангием называют клетку, в которой образуются подвижные половые элементы — гаметы.
Споры растений — микроскопические зачатки некоторых растений, имеющие разное происхождение, служащие для их размножения и (или) сохранения при неблагоприятных условиях. Представляют собой одноклеточные, реже двуклеточные или состоящие из нескольких клеток образования. Обычно более или менее шарообразной, эллипсоидальной формы, реже — цилиндрической и др.
Упоминания в литературе
Органы полового размножения – антеридии (муж-ские) и архегонии (женские) чаще располагаются на растении группами, обычно окружены листовидными выростами или другими защитными образованиями. Оплодотворение яйцеклетки подвижными двухжгутиковыми сперматозоидами, образующимися в антеридиях, возможно только при наличии капельно-жидкой воды. Слияние гамет и развитие зиготы происходит внутри архегония. Из зиготы в течение определённого времени (от нескольких месяцев до 2 лет) развивается многоклеточный диплоидный спорофит (специализированный орган размножения), называемый спорогоном. Он состоит из верхней спороносной части (коробочки) и нижней – ножки со стопой, врастающей в ткань гаметофита . Из образующихся путём редукционного деления спор развивается многоклеточное ветвистое нитчатое или пластинчатое образование – протонема, на которой закладываются почки, дающие начало пластинчатым талломам или облиственным побегам – гаметофорам. Массовое участие мхов в растительном покрове оказывает существенное воздействие на среду обитания других растений и животных. В районах повышенного увлажнения в умеренных зонах накапливаются значительные (толщиной до 11 м) залежи торфа с преобладанием мхов.
Тычинка состоит из тычиночной нити и пыльника. Основанием тычиночной нити тычинка прикреплена к цветоложу. Пыльник имеет две половинки (теки), соединенные связником, который является продолжением тычиночной нити. Каждая из тек включает два гнезда (пыльцевые мешки), где образуются микроспоры, а затем пыльца. В пыльнике происходят два важнейших процесса: микроспорогенез – образование микроспор и микрогаметогенез – развитие из микроспор мужского гаметофита , или пыльцевого зерна.
Связанные понятия (продолжение)
Размноже́ние расте́ний — совокупность процессов, приводящих к увеличению числа особей некоторого вида; у растений имеет место бесполое, половое и вегетативное (бесполое и половое размножение объединяют в понятие генеративное размножение). Изучение различных аспектов размножения является предметом репродуктивной биологии.
Талло́м, или слоеви́ще (от др.-греч. θαλλός — молодая, зелёная ветвь), — ботанический термин, применяемый для обозначения одноклеточного, многоклеточного или не дифференцированного на клетки (многоядерного) тела водорослей, грибов, лишайников, а также антоцеротовых и некоторых печёночных мхов.
Плаунови́дные (лат. Lycopodióphyta) — отдел высших споровых растений. Являются древнейшими сохранившимися до наших дней сосудистыми растениями. Произошли от риниофитовидных в середине девонского периода, достигнув рассвета в каменноугольном периоде. На данный момент все древесные формы плауновидных являются вымершими, а всего осталось около одной тысячи видов.
Зооспо́ра (др.-греч. ζῷον — животное и σπορά — посев, семя), или зоогони́дий, или бродя́жка — стадия жизненного цикла многих водорослей и некоторых низших грибов. Представляют собой жгутиконосцев, перемещающихся в жидкой среде с помощью биения одного или нескольких жгутиков. Многие водоросли на этой стадии обладают хроматофором, стигмой и сократительными вакуолями. Зооспоры некоторых желто-зелёных водорослей обладают многочисленными ядрами и несколькими парами жгутиков (синзооспоры).
Апика́льная меристе́ма — группа меристематических (образовательных) клеток, организованных в ростовой центр, занимающая терминальное положение в стебле и обеспечивающая образование всех органов и первичных тканей побега.
Спо́ры (греч. σπορά, σπόρος — сев, посев, семя) — клетки растений и грибов, служащие для их размножения и рассеяния.
Ризо́иды (от др.-греч. ῥίζα — корень и εἶδος — вид) — нитевидные образования из одной или нескольких однорядных клеток; служат для прикрепления к субстрату и поглощения из него воды и питательных веществ. Имеются у мхов, лишайников, некоторых водорослей и грибов (например, Rhizopus). Внешне напоминают корневые волоски.
Спо́ры (от др.-греч. σπορά — сеяние, посев) — общий термин для репродуктивных структур грибов (Fungi). Споры грибов возникают в результате бесполого размножения или полового процесса и служат для размножения.
Половое размножение — процесс у большинства эукариот, связанный с развитием новых организмов из половых клеток (у одноклеточных эукариот при конъюгации функции половых клеток выполняют половые ядра).
Эпиде́рма (эпиде́рмис, ко́жица) — внешняя первичная покровная ткань растений, обычно однослойная, покрывающая молодые стебли и остальные наземные органы (листья, лепестки, плоды и др.). Представляет собой наружный слой клеток, образующийся из протодермы конуса нарастания.
Спорофилл, спороносный лист, споролистик — боковой орган побега спорофита сосудистых растений, более или менее видоизменённый лист, на котором располагается один или несколько спорангиев.
Бесполое размножение, или агамогенез — один из способов размножения, при котором следующее поколение развивается из соматических клеток без участия репродуктивных клеток — гамет. Следует отличать бесполое размножение от однополого размножения (партеногенеза), который является особой формой полового размножения.
Оогамия (от др.-греч. ᾠόν — яйцо и γάμος — брак) — это вид полового процесса, при котором сливаются резко отличающиеся друг от друга половые клетки — крупная неподвижная яйцеклетка с мелкой, обычно подвижной, мужской половой клеткой (сперматозоидом или спермием).
Интегуме́нт (от лат. integumentum — покрывало, покров) — термин, служащий в биологии для обозначения покрова (оболочки) организма или его части. Широко применим в ботанике. Иногда, преимущественно по отношению к членистоногим, используется в зоологии.
Пло́идность — число одинаковых наборов хромосом, находящихся в ядре клетки или в ядрах клеток многоклеточного организма.
Флоэ́ма (от греч. φλοῦς — кора) — то же, что и луб — проводящая ткань сосудистых растений, по которой происходит транспорт продуктов фотосинтеза к частям растения, в которых он не происходит: подземные части, конусы нарастания, цветки, плоды и др. Вместе с ксилемой (древесиной), обеспечивающей транспорт воды и минеральных солей, образует проводящие пучки.
Антоцеротови́дные, или Антоцеротофи́ты, или Антоцеро́товые мхи (лат. Anthocerotophyta) — отдел мохообразных растений, для которых характерны пластинчатые талломы. Ранее Антоцеротовидные (Антоцеротовые) рассматривались как класс отдела Моховидные, или Мхи (Bryophyta) — Anthocerotopsida.
Ксиле́ма (от греч. ξύλον — древеси́на) — основная водопроводящая ткань наземных сосудистых растений; один из двух подтипов проводящей ткани растений, наряду с флоэмой — лубом.
Неолектомице́ты (лат. Neolectomycetes) — класс грибов отдела Аскомицеты (Ascomycota), один из 4 классов, относящихся к подотделу Taphrinomycotina. Класс монотипный, к нему относятся также монотипные порядок и семейство, соответственно, Neolectales и Neolectaceae, и род Neolecta с четырьмя видами.
Голосеменны́е расте́ния (лат. Gymnospérmae) — древняя группа семенных растений, появившаяся в верхнем девоне, около 370 млн лет назад.
Печёночные мхи, или Печёночники, или Маршанциевые мхи (лат. Marchantiophyta ), — отдел растений со слабо развитой протонемой; распространены по всему земному шару, особенно в тропиках. Число видов — от шести до восьми тысяч.
Разделение полов у растений — явление, при котором у растений одного вида имеются как мужские (тычиночные), так и женские (пестичные) цветки. Такая особенностью растений является приспособлением, препятствующим самоопылению и способствующим перекрёстному опылению.
Зиго́та (от др.-греч. ζυγωτός — удвоенный) — диплоидная (содержащая полный двойной набор хромосом) клетка, образующаяся в результате оплодотворения (слияния яйцеклетки и сперматозоида). Зигота является тотипотентной клеткой, то есть способной породить любую другую. Термин ввёл немецкий ботаник Э. Страсбургер.
Зигомико́та (лат. Zygomycota) — отдел грибов, объединяющий 10 порядков, 27 семейств, около 170 родов и более 1000 видов. Отличаются развитым ценоцитным мицелием непостоянной толщины, в котором септы образуются только для отделения репродуктивных органов.
Вегетати́вное размноже́ние — образование новой особи из многоклеточной части тела родительской особи, один из способов бесполого размножения, свойственный многоклеточным организмам. У водорослей и грибов происходит путём отделения неспециализированных участков таллома или посредством образования специализированных участков (выводковые почки водоросли сфацелярии и др.).
Полово́й проце́сс, или оплодотворе́ние, или амфими́ксис (др.-греч. ἀμφι- — приставка со значением обоюдности, двойственности и μῖξις — смешение), или сингамия — процесс слияния гаплоидных половых клеток, или гамет, приводящий к образованию диплоидной клетки зиготы. Не следует смешивать это понятие с половым актом (встречей половых партнёров у многоклеточных животных).
Периде́рма (от греч. περι — около и греч. δερμα — кожа) — комплекс тканей, состоящий из феллогена и его производных — феллодермы (откладывается внутрь) и феллемы, или пробки (вторичной покровной ткани, откладывается наружу). Перидерма является одним из чётко выраженных конструктивных элементов строения стебля высших растений, которые невозможно отнести ни к тканям, ни к органам. Такие элементы называются анатомо-топографическими зонами.
Мицелий (грибница) — вегетативное тело грибов и актиномицетов (некоторые исследователи, подчёркивая бактериальную природу актиномицетов, называют их аналог грибного мицелия тонкими нитями), состоящее из тонких (1,5—10 мкм толщиной у грибов и 0,5—1,0 мкм у актиномицетов) разветвлённых нитей, называемых гифами. Развивается в субстрате и на его поверхности.
Субстрат (от лат. substratum — основа, подстилка) — в биологии место обитания и развития организмов. Субстраты служат местом прикрепления животного, растения или другого организма и могут выполнять роль питательной среды.
Хламидоспо́ры, или ге́ммы, — органы вегетативного размножения некоторых зигомицетовых и других грибов, представляющие собой толстостенные увеличенные клетки округлой формы, также используемые для переживания неблагоприятных условий.
Пиреноид — особое включение (дифференцированная область) внутри хлоропласта. Встречаются у некоторых групп эукариотических водорослей и у антоцеротовых мхов. Функционально представляет собой место запасания фермента рибулозобисфосфаткарбоксилаза (рубиско) и/или центр синтеза сахаров при помощи этого фермента. Вокруг пиреноида у зелёных водорослей откладываются запасные продукты (крахмал).
Зародышевый мешок (англ. female gametophyte, embryo sac) — женский гаметофит, половое поколение покрытосеменных растений.
Стро́билы (лат. strobilus от др.-греч. στρόβῑλος — кубарь, сосновая или еловая шишка) — видоизменённые побеги или части побегов, несущие на себе спорангии.
Хитридиомице́ты (лат. Chytridiomycota) — отдел царства грибов (Fungi). Объединяет более 120 родов и около 1000 видов. Мицелий слабо развит, основная масса таллома представляет собой т. н. плазмодий, из которого вырастают ризоидные гифы. Самые примитивные представители совершенно не имеют мицелия, и тело их в вегетативном состоянии представлено одиночной клеткой, иногда лишённой жёсткой клеточной стенки. Основа клеточной стенки хитиново-глюкановая, как и у высших грибов.
Гастеромице́ты, гастромице́ты, нутревики́ — полифилетическая группа таксонов грибов класса базидиомицетов. К гастеромицетам относят около 1000 видов из 110 родов.
Вегетати́вные о́рганы растений (от лат. vegetativus — растительный) — части растения, выполняющие основные функции питания и обмена веществ с внешней средой.
Во́львокс (лат. Volvox) — род подвижных колониальных организмов, относящийся к отделу зелёных водорослей. Обитают в стоячих пресных водоёмах. При массовом размножении вызывают цветение воды, окрашивая её в зелёный цвет.
Высшие споровые растения — неформальный термин, объединяющий высшие растения, размножающиеся и распространяющиеся главным образом спорами. Эта группа таксонов обычно противопоставляется семенным растениям, которые для размножения используют не споры, а семена. Высшие споровые признаются одними из первых наземных растений.
Спешу обрадовать, мы добрались до изучения семенных растений! К ним относятся голосеменные и покрытосеменные (цветковые). До этого размножение шло только с помощью спор: у мхов, папоротников, хвощей и плаунов - высших споровых растений. Настало время открыть новую интересную главу этой книги, посвященную растениям, которые размножаются с помощью удивительного изобретения природы - семени.
Голосеменные - распространенная древняя группа растений, включающая небольшое число видов. Главной особенностью данной группы являются "голо" (то есть открыто) лежащие семяпочки и, в дальнейшем, развивающиеся из них семена. Иными словами, у голосеменных растений отсутствуют замкнутые вместилища для семян.

На примере типичного представителя - сосны обыкновенной, относящейся к классу хвойных, поговорим о характерных чертах данного класса и голосеменных растений в целом.
Общие признаки
Все голосеменные представлены древесными формами: деревьями и кустарниками. Травы отсутствуют.
Хвоинки (хвоя) - игольчатые видоизменения листьев. Сохраняются долгие годы, у некоторых сосен до 45 лет. Хвоя лиственниц опадает ежегодно.

Древесина голосеменных обладает большим запасом механической прочности. Это связано с ее особенностями: она состоит из трахеид с окаймленными порами, паренхима развита слабо. Либриформ (древесные волокна) и настоящие сосуды отсутствуют (исключение - гнетовые, имеют сосуды). Клетки-спутницы во флоэме также отсутствуют.
В древесине и коре имеются каналы, заполненные смолой. Однако, есть исключения - у гинкго смола не образуется вовсе.
Несколько веков назад в России целенаправленно создавались и охранялись, так называемые, корабельные рощи. Это, прежде всего, требовалось для флота, так как мачты кораблей изготавливали из сосен, отвечающих всем требованиям - корабельных (гладкий, твердый и прочный прямой ствол с минимальным количеством сучков и смолы).

Семяпочки и развивающиеся из них семена лежат "голо", открыто, для них нет закрытых вместилищ, отсутствует завязь. В сравнении с высшими споровыми растениями, размножение семенами ставит голосеменных на более высокий уровень организации.
Голосеменным растениям для размножения не требуется вода, опыление у них происходит с помощью ветра. Этот процесс перестал быть зависимым от капельно-жидкой среды, как было у мхов и у папоротников. Благодаря этому голосеменные получили большое преимущество и смогли расселиться по всей Земле, в том числе в засушливых районах. Они господствовали в юрском периоде, когда климат стал более сухим и жарким.

Обитают голосеменные в местах с холодным климатом и достаточным количеством влаги. Имеются виды, обитающие в жарких странах: растение вельвичия удивительная обитает в пустынях южной Африки.
Строение и жизненный цикл
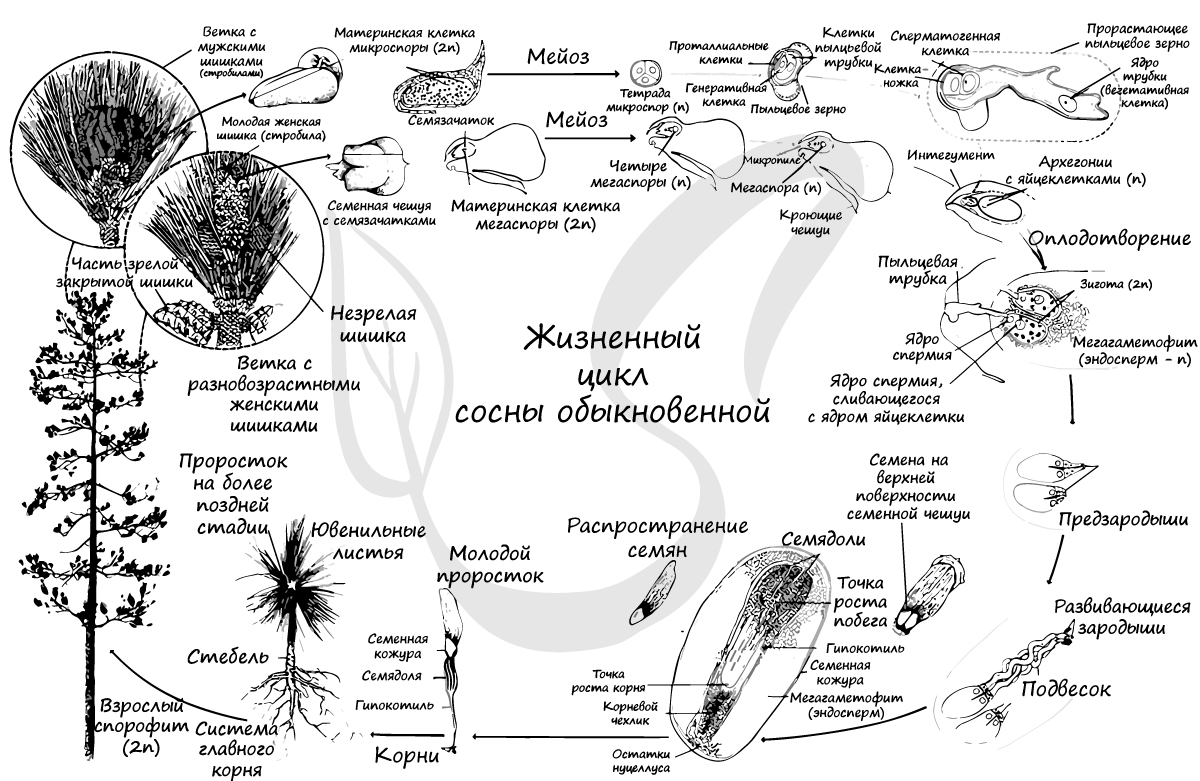
Жизненный цикл голосеменных состоит из чередования бесполого поколения - спорофита (диплоиден, 2n), и полового поколения - гаметофита (гаплоиден, n). Господствует (доминирует) в цикле спорофит (2n) - это взрослое растение сосны.
Голосеменные относятся к разноспоровым, как и все семенные растения. Они образуют разные споры: крупные женские (мегаспоры) и мелкие мужские (микроспоры). Образуются они в спорангиях, расположенных на спорофиллах, которые собраны в стробилы (шишки) - от лат. strobilus - сосновая шишка.
Мужские шишки (стробилы)
К концу весны у основания молодых побегов образуются мужские шишки (стробилы) - мелкие, собранные в тесные группы, желтого цвета. Чешуи мужских шишек представляют собой микроспорофиллы. Микроспорофиллы - гомологи тычинок, которые крепятся к оси каждой шишки спирально, с нижней стороны, и имеют два пыльцевых мешка - микроспорангия.
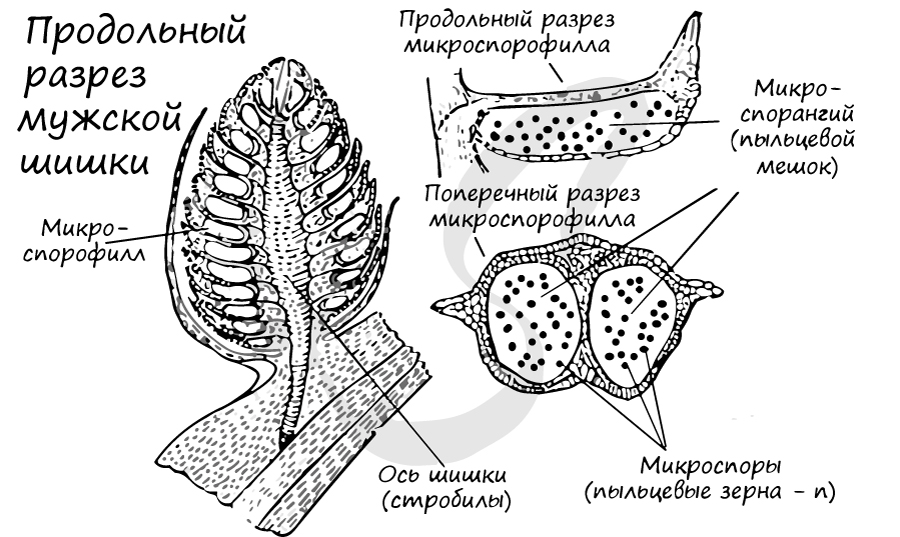
Образование мужского гаметофита
Из материнских клеток (2n) в микроспорангии путем мейоза образуются 4 микроспоры (n). Строение микроспоры следующее: она покрыта экзиной (от гр.exo снаружи, вне) - наружная оболочка, изнутри интиной (от лат. intus внутри) - внутренней оболочкой. В составе микроспоры имеются также два воздухоносных мешка, образованных в результате отслоения экзины от интины и возникновения полости между ними.
Микроспора делится, не покидая спорангия, преобразуется в заросток. При делении из ядра микроспоры образуются две клетки. Одна из них превращается в две заростковые клетки (протоллиальные - от греч. проталлиум - заросток) - быстро отмирают и исчезают. Их функция до конца не изучена.
Из другой клетки в ходе митоза также образуются две: антеридиальная, из которой развиваются мужские половые клетки - спермии (неподвижные, без жгутиков в отличие от сперматозоидов), и более крупная вегетативная клетка, из которой в дальнейшем формируется пыльцевая трубка.
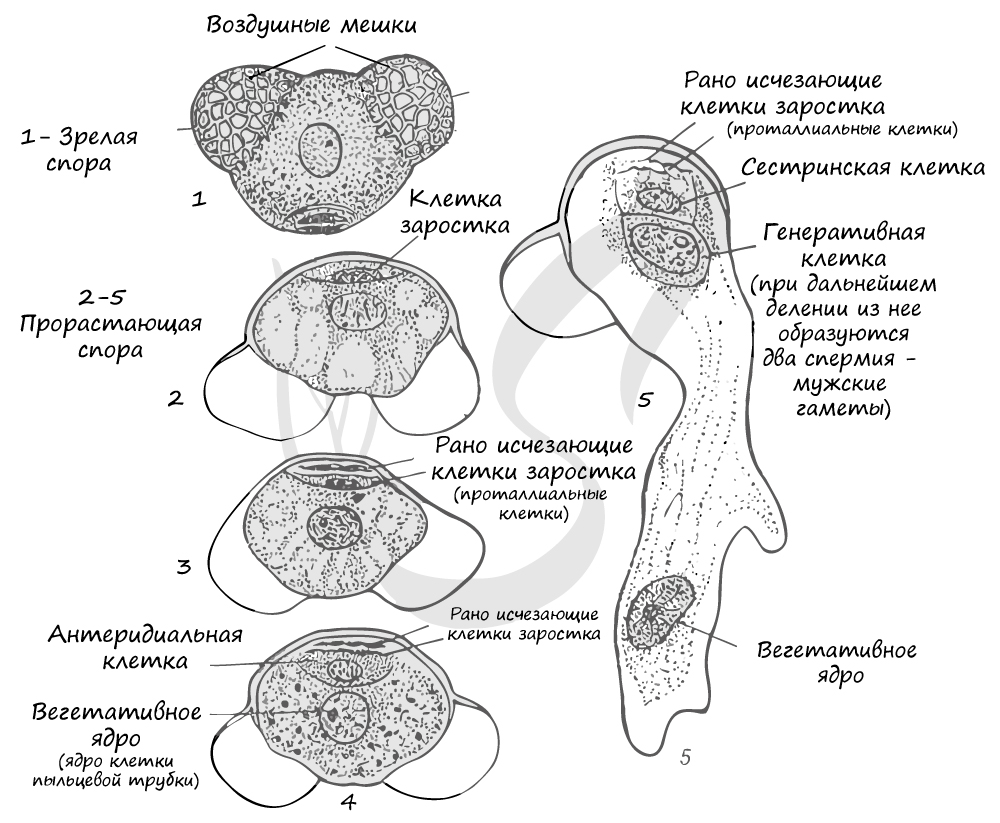
Мужской гаметофит сильно упрощен, антеридии отсутствуют. Формируется он прямо внутри микроспоры, которая в итоге превращается в пыльцевое зерно. Совокупность пыльцевых зерен называется пыльца.
При вскрытии (нарушении целостности) микроспорангия, или пыльцевого мешка, пыльца высыпается во внешнюю среду и достигает женской шишки, где, в результате опыления, внутри семязачатка происходит дальнейшее развитие мужского гаметофита.
Образование женского гаметофита
На тех же самых соснах, где расположены мужские шишки, лежат и женские. Весной на верхушке молодого побега появляются мелкие (около 5 мм) красноватые шишки - это женские шишки (стробилы). Состоят они из оси (стержня) , на котором располагаются две чешуи: кроющая и семенная. На верхней стороне у основания семенной чешуи лежат два семязачатка.
Кроющая чешуя представляет собой видоизмененный лист, в его пазухе находится семенная чешуя. Семенная чешуя - видоизмененный боковой побег.
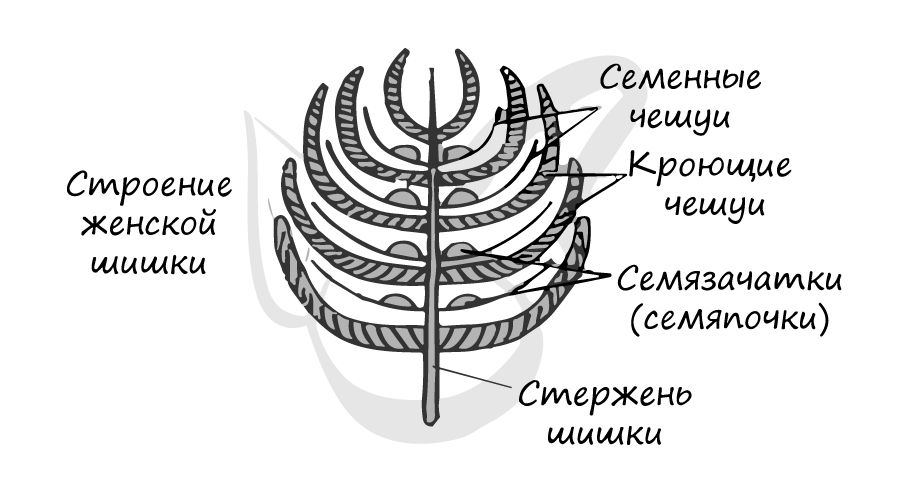
Женские шишки (стробилы)
Именно открыто расположенные семязачатки (семяпочки) служат причиной, по которой этот отдел растений называется - голосеменные.
В женских шишках, в отличие от мужских, каждая чешуя гомологична целой мужской шишке (стробилу). То есть одна чешуя - целой мужской шишке, а не отдельным ее микроспорофиллам (чешуям)!
Молодой семязачаток состоит из нуцеллуса, интегумента и фуникулуса. Нуцеллус (от лат. nucella - орешек) - центральная часть семяпочки, соответствующая мегаспорангию. Интегумент (от лат. integumentum покрывало) - покров семяпочки, вырастающий из ее центральной части - нуцеллуса. В зрелом семени интегумент преобразуется в семенную кожуру. Фуникулус (от лат. funiculus канатик, верёвка) или семяножка - часть семязачатка, соединяющая его с мегаспорофиллом (семенным чешуями).
На интегументе около вершины располагается микропиле (пыльцевход) - через него после опыления пыльцевая трубка проникает в нуцеллус. Между интегументом и нуцеллусом имеется густая жидкость, выступающая из микропиле. Подсыхая, она втягивается внутрь семязачатка и затягивает вместе с собой пыльцу, осевшую на ней.
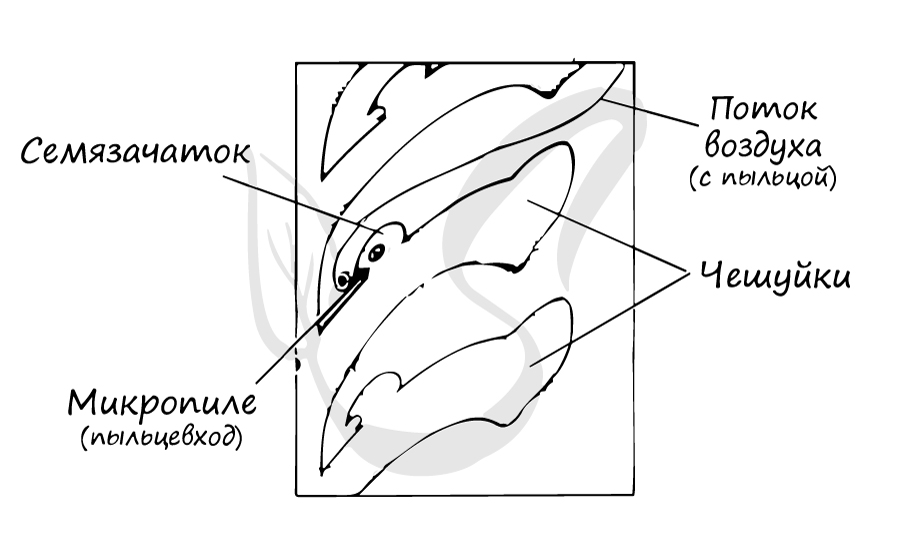
Образование женского гаметофита
В средней части обособляется спорогенная клетка (2n) (археспориальная - от греч. arche начало и sporá семя). В результате ее митотического деления образуются материнские клетки спор - спороциты (2n), однако и сама археспориальная клетка может выступать в роле спороцита, минуя стадию митоза. Спороциты (2n) делятся мейозом на четыре гаплоидные (n) мегаспоры.
Три мегаспоры отмирают, остается одна, которая многократно делится митозом и формирует эндосперм - запасное питательное вещество. Обратите на этот факт особое внимание: у голосеменных эндосперм гаплоидный (n) и образуется до оплодотворения. Такой эндосперм называется - первичный, он соответствует женскому гаметофиту.
Как и мужской, женский гаметофит весьма упрощен и заключен внутри мегаспоры. На верхушке женского гаметофита (мегагаметофита) образуется архегоний с яйцеклеткой (n). У гнетовых архегонии отсутствуют.
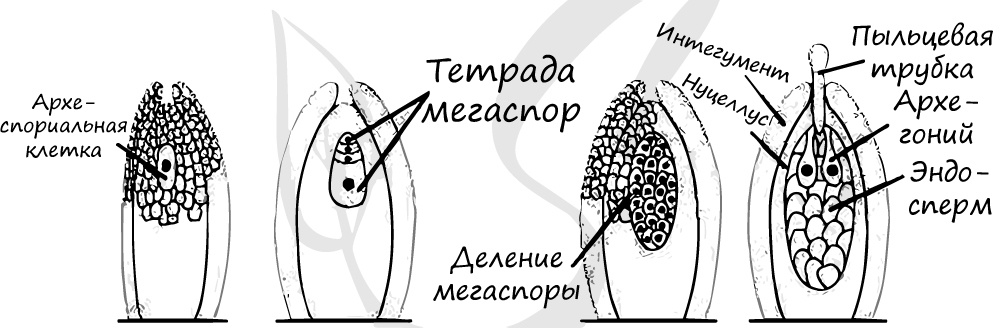
Жизненный цикл
На спорофите (2n) в микроспорангиях из материнских клеток (2n) путем мейоза образуются микроспоры (n). Из микроспоры формируется пыльцевое зерно. Пыльца (пыльцевые зерна (n)) с помощью ветра попадает в женские шишки, где улавливается густой жидкостью между интегументом и нуцеллусом, выступающей из микропиле. Жидкость засасывает пыльцу внутрь семязачатка на нуцеллус (в пыльцевую камеру). После того, как опыление произошло, микропиле зарастает. Чешуи шишки смыкаются и склеиваются смолой.
Семязачатки в этот момент еще не готовы к оплодотворению, так что от момента опыления до оплодотворения проходит около 13 месяцев. За это время в семязачатке формируется эндосперм, женская шишка увеличивается до 3-4 см и приобретает зеленую окраску.
Оказавшись на мегаспорангии, наружная оболочка пыльцевого зерна (экзина) разрывается, из вегетативной клетки в направлении архегония начинает расти пыльцевая трубка. Антеридиальная клетка делится на генеративную (спермагенную) и клетку-ножку антеридия (функция последней до сих пор не изучена). Спермагенная клетка попадает в пыльцевую трубку, а из нее - в архегоний.
Непосредственно перед оплодотворением спермагенная клетка делится на два спермия (n), один из которых отмирает, а другой сливается с яйцеклеткой (n). Образуется зигота (2n), из которой формируется и растет зародыш благодаря эндосперму - запасу питательных веществ.
Окончательно созревают семена к осени на второй год после опыления, к этому моменту женские шишки увеличиваются в размерах до 6 см. Зеленая окраска меняется на серую, чешуйки расходятся, и семена, образовавшиеся из семязачатков, высыпаются. Из семени прорастает взрослое растение - спорофит (2n). Цикл замыкается.
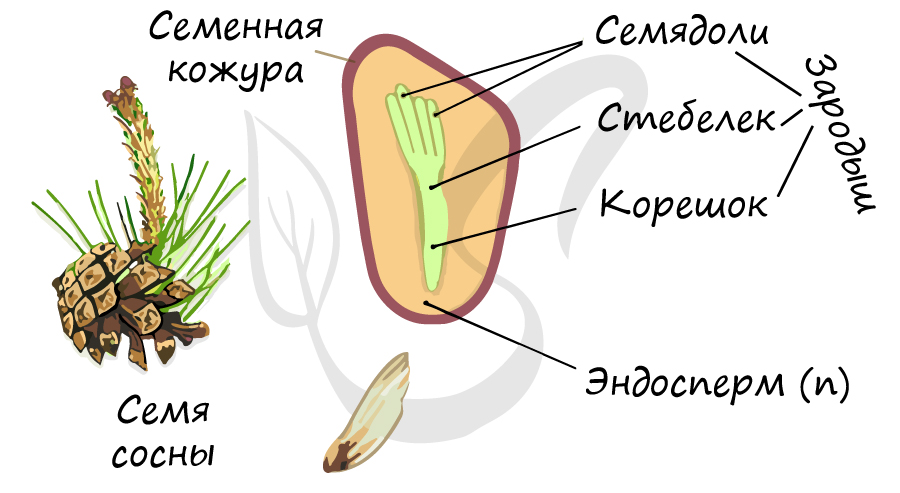
Строение семени
Семенная кожура, защищающая семя от пересыхания и неблагоприятных факторов внешней среды, образована разросшимся интегументом.
Зародыш (2n) формируется в результате митотического деления образовавшейся зиготы. Состоит из зародышевого корешка, стебелька и почечки.
Число семядолей у голосеменных различается - от 2 до 15. Семядоли имеют доступ к запасным питательным вещества (эндосперму).
Запасные питательные вещества накапливаются в эндосперме (n). Особенностью в строении семени голосеменных, по сравнению с семенем покрытосеменных (цветковых) является наличие гаплоидного эндосперма (n). Не забывайте, что эндосперм у голосеменных это производное мегагаметофита (n), исходя из этого становится понятно, почему ткань гаплоидна. У цветковых, в отличие от голосеменных, эндосперм триплоиден (3n).
Фитонциды
Фитонциды (от греч. phyton - растение и лат. caedo - убиваю) - образуемые растениями, биологически активные вещества, убивающие или приостанавливающие размножение других организмов, главным образом - микробов. Обычно выделяются растениями в газообразном виде, к примеру, аллицин у лука и чеснока. Наличие фитонцидов играет крайне важную роль в формировании устойчивости растения к грибным заболеваниям.
Фитонциды имеют медицинское значение, из них изготавливаются некоторые препараты. За лето гектар лиственного леса выделят 2 кг фитонцидов, хвойного - 5 кг, можжевельника - 30 кг! Санатории часто располагаются в сосновых борах, где наблюдается повышенная концентрация фитонцидов. Вдыхание такого воздуха очень полезно при заболеваниях дыхательной системы инфекционной природы (когда возбудителями являются бактерии, грибы).

Значение голосеменных
Трудно переоценить значение голосеменных для человека, они очень важны. Голосеменные - источники высококачественной древесины, продуктов ее переработки. Являются звеном в цепи питания (продуцентами), основой многих биоценозов. Хвойные растения в больших количествах выделяют фитонциды, имеющие медицинское значение. Из смолы хвойных получают канифоль, скипидар, лаки. Кедровых орехи - это семена нескольких видов растений из рода сосна, которые употребляют в пищу.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: