
1958 ф стьюард выращивает целое растение моркови из единственной клетки
Добавил пользователь Алексей Ф. Обновлено: 20.09.2024
В 1880-х годах Аугуст Вайсман (August Weismann), профессор зоологии и сравнительной анатомии Фрайбургского университета (University of Freiburg), предположил, что генетическая информация постепенно сокращается при каждом делении клетки.
Теорию Вайсмана экспериментально подтвердил немецкий эмбриолог Вильгельм Ру (Wilhelm Roux). Когда он при помощи горячей иглы уничтожил одну из клеток двухклеточного эмбриона лягушки, развилась только половина зародыша.
В 1894 году немецкий биолог Ханс Дриш (Hans Driesch) оспорил теорию Вайсмана и Ру, экспериментально выявив, что бластомеры (клетки, образованные в результате дробления зиготы), изолированные из двух- и четырехклеточных эмбрионов морского ежа, способны развиться в небольшие личинки.
В 1901 году немецкий ученый Ханс Шпеман (Hans Spemann) разделил двухклеточный зародыш саламандры на две части, каждая из которых развилась в полноценный организм. Результат эксперимента подтвердил, что на ранней стадии развития эмбриональные клетки содержат достаточную для создания самостоятельного организма информацию.
В 1902 году американский генетик Уолтер Саттон (Walter Sutton) выдвинул гипотезу, что генетическая информация хромосомы содержится в клеточном ядре.
В 1914 году Ханс Шпеман провел первые опыты по пересадке ядра из одной клетки в другую.
В 1938 году Шпеман предположил, что существует возможность пересадки ядра одной клетки в безъядерную яйцеклетку.
Так была заложена теоретическая основа последующих экспериментов по клонированию.
В 1962 году Джон Гердон (John Gurdon), профессор Оксфордского университета, заявил, что клонировал южно-африканскую лягушку. Ему удалось добиться развития яйцеклеток лягушки с чужим ядром до достаточно поздних стадий, примерно в двух процентах случаев особи превращались во взрослых лягушек. В 2012 году Гердон за свои исследования стал лауреатом Нобелевской премии по медицине или физиологии.
В этом же году китайский исследователь Тун Дичжоу (Tong Dizhou) впервые клонировал рыбу. Он пересадил генетический материал взрослого азиатского карпа в икринку, из которой появилась новая особь, которая впоследствии принесла потомство.
В 1964 году профессор Корнелльского университета Фредерик Стюард (Frederic Steward) вырастил целую морковь из полностью изолированных клеток корневища, доказав таким образом возможность клонирования с использованием дифференцированных (изолированных) клеток.
В 1979 году Карл Илменси (Karl Illmensee) заявил, что ему удалось клонировать трех мышей.
В 1983 году Кэри Муллис (Kary Mullis) разработал метод полимеразной цепной реакции (ПЦР), позволяющий добиться значительного увеличения малых концентраций определенных фрагментов нуклеиновой кислоты (ДНК) в биологическом материале (пробе).
В 1984 году датский ученый Стин Вилладсен (Steen Willadsen) клонировал овцу из эмбриональных клеток. Это был первый эксперимент по клонированию млекопитающего. Вилладсен использовал метод пересадки ядра (nuclear transfer).
В 1986 году Вилладсен клонировал корову из дифференцированных клеток однонедельного эмбриона.
В этом же году профессора Университета Висконсина Нил Ферст (Neal First), Рэндал Прэзер (Randal Prather) и Уиллард Айстоун (Willard Eyestone) также клонировали корову из клеток зародыша.
В 1995 году профессора Рослинского института в Шотландии Иэн Уилмут (Ian Wilmut) и Кит Кэмпбелл (Keith Campbell) с помощью генетического материала двух изолированных эмбрионов успешно клонировали двух овец – Меган и Мораг.
В 1996 году Уилмут и Кэмпбелл впервые поставили эксперимент по клонированию животного из клеток взрослой особи, в результате которого 5 июля 1996 года на свет появилась овца Долли.
27 февраля 1997 года в английском журнале Nature была опубликована сенсационная статья эмбриолога и генетика Иэна Уилмата об успешном клонировании овечки Долли.
В 1997 году в лаборатории Дона Вольфа (Don Wolf) Орегонского регионального центра по изучению приматов (Oregon Regional Primate Research Center) ученым удалось клонировать двух макак-резусов.
В этом же году президент США Билл Клинтон запретил использовать государственные средства для финансирования работ по клонированию человека. (На самом деле таких работ никогда не проводилось, а запрещено было выделять госсредства на получение новых линий человеческих эмбриональных стволовых клеток. За государственный счет разрешалось продолжать эксперименты только с небольшим числом имевшихся на тот момент линий ЧЭСК – ВМ.)
В 1997 году Иэн Вилмут и Кит Кэмпбелл использовали выращенные в лабораторных условиях клетки кожи с генетически вживленным в них человеческим геном и клонировали еще одну овцу – Полли.
В 1998 году Долли родила трех здоровых ягнят, зачатых естественным путем.
В этом же году в Университете штата Гавайи группа ученых во главе с профессором Риузо Янагимачи (Ryuzo Yanagimachi) клонировала 50 мышей из клеток взрослых особей. Первая мышь-клон получила прозвище Кумулина.
5 марта 2000 года английские ученые провели первую успешную операцию по клонированию свиней.
22 декабря 2001 года в Техасском университете агрокультуры и машиностроения (Texas A&M University) родилась первая в мире клонированная домашняя кошка по кличке Сиси (CopyCat, CC). Через два года в университете родились первый клонированный олень – Дьюи и первая клонированная лошадь – Прометея.
В 2001 году ученые из компании Advanced Cell Technology, Inc. объявили о рождении быка-гаура по кличке Ноа, который стал первым клонированным животным, относящимся к вымирающим видам. Этот эксперимент открыл перспективу спасения исчезающих видов животных путем клонирования.
В 2003 году знаменитую на весь мир овцу Долли усыпили. Причиной послужил прогрессирующий рак легких, вызванный вирусом. Долли было 6,5 лет.
В 2005 году в Южной Корее на свет появилась первая в мире клонированная собака – африканская борзая Снаппи.
В июне 2013 года группа ученых японского научного центра RIKEN сумела клонировать мышь, используя лейкоциты, выделенные из капли периферической крови животного. Ранее лейкоциты не использовались для клонирования.
Несмотря на ряд заявлений о проведении успешных опытов по клонированию человеческого эмбриона (1998, 2004 – в Южной Корее, 2002 – в США), научного подтверждения этому не было.
В 2013 году в США выходец из Казахстана Шукрат Миталипов смог извлечь стволовые клетки из эмбрионов, выращенных из человеческих клеток кожи. Научный коллектив под руководством профессора Орегонского университета науки и здоровья (Oregon Health and Science University) Миталипова извлек ДНК из донорской яйцеклетки, полученной от молодой женщины, поместил в нее генетическую информацию из клетки кожи и заставил клетку делиться, но не путем стимулирования электрическими разрядами, как в случае с клетками овечки Долли, а с помощью кофеина. Полученный эмбрион развивался 5-6 дней и достиг размера в 120 клеток, после чего Миталипову удалось извлечь из него живые стволовые клетки.
По мнению британского ученого Иэна Уилмута – создателя овечки Долли, существующие процедуры клонирования очень неэффективны.
Овечка Долли была единственным выжившим из 277 эмбрионов. Последующие исследования показали, что в лучшем случае успехом заканчиваются лишь 5% попыток.
Запрет на проведение опытов по репродуктивному клонированию человека действует в Австралии, Австрии, Аргентине, Бельгии, Бразилии, Канаде, Чехии, Дании, Финляндии, Франции, ФРГ, Италии, Японии, Швеции, Швейцарии и других странах.
В 2002 году в России был введен пятилетний мораторий на клонирование человека, срок действия которого истек в 2007 году.
В 2010 году Госдума РФ приняла закон о продлении моратория. Закон при этом не запрещает клонирование клеток и организмов в научно-исследовательских целях, выращивание органов для трансплантации и клонирование животных.
Матричные процессы составляют основу способности живых организмов к воспроизведению. В клеточном ядре происходит удвоение ДНК. Новая молекула воспроизводится на матрице старой и представляет собой ее точную копию. Информационная РНК синтезируется на матрице ДНК в виде точной копии одного из участков ДНК. Далее на матрице иРНК происходит синтез белков.
Трансляция. Перевод последовательности нуклеотидов иРНК в последовательность аминокислот синтезируемых белков называют трансляцией.
В активном центре рибосомы размещаются два триплета иРНК и соответственно две тРНК. Рибосома перемещается по иРНК не плавно, а прерывисто, триплет за триплетом. На каждом шаге присоединяется новая аминокислота. Транспортные РНК перемещаются со своей аминокислотой к рибосоме и "примеряют" свой антикодон к очередному кодону иРНК, находящемуся в активном центре (комплементарные нуклеотиды хорошо подходят друг другу, между ними возникают водородные связи). Если антикодон оказывается не комплементарным, то тРНК удаляется в цитоплазму к другим рибосомам. Если же он оказывается комплементарным, то тРНК присоединяется к кодону водородными связями.
Далее особый фермент рибосомы подсоединяет синтезируемую полипептидную цепь к "прибывшей" аминокислоте. Ее транспортная РНК продолжает удерживать всю формирующуюся белковую цепь на кодоне иРНК до прихода следующей тРНК. Освободившаяся тРНК перемещается в цитоплазму "на поиск" аминокислот. Рибосома перескакивает на следующий триплет иРНК, и процесс повторяется. Интервал между перескакиваниями продолжается не более 1/5 — 1/6 с, а вся трансляция среднего белка — 1-2 минуты.
На одной нити иРНК "трудятся" сразу несколько рибосом. На иРНК, содержащей информацию о белке гемоглобине, размещается до 5 рибосом, на некоторых других иРНК — до 20 рибосом. Когда в активном центре рибосомы оказывается один из трех триплетов, кодирующих знаки препинания между генами, синтез белка завершается.
Освободившаяся рибосома отправляется на другую иРНК. Рибосомы универсальны и могут синтезировать полипептиды по любой матрице иРНК. Субъединицы рибосом соединяются только для синтеза белка, после окончания синтеза они вновь разъединяются.
Сворачивание в спираль и приобретение третичной структуры происходит по мере синтеза белковой цепи, поэтому зачастую невозможно восстановление денатурированных белков со сложной пространственной структурой, даже если их первичная структура сохранена. Сворачивание белковой цепи целой молекулы происходит по-иному, и белок оказывается неактивным.
Многие белки — например, пищеварительные ферменты — очень активны и способны переварить саму клетку, поэтому синтезируемые молекулы белков сразу попадают в эндоплазматическую сеть (к мембране которой прикреплены синтезирующие белки рибосомы) и по ее каналам перемещаются к комплексу Гольджи, а от него в тот участок клетки или в ту часть организма, где требуется этот вид белков. Синтез небелковых соединений клетка осуществляет в два этапа. Сначала рибосомы производят трансляцию специфического белка-фермента. Затем при его участии образуется молекула необходимого соединения — углевода, жиров и т.д. Сходным образом синтезируются и другие соединения: витамины, небелковые гормоны и пр.
Процесс синтеза белков требует затрат большого количества специфической энергии АТФ, которую нельзя заменить иным источником. Только на присоединение каждой аминокислоты к тРНК расходуется энергия одной молекулы АТФ, а в среднем белке 400-500 аминокислот. Общее количество затрачиваемой энергии существенно превышает суммарную химическую энергию образующихся пептидных связей между молекулами аминокислот. По этой причине белки редко используются клеткой в качестве топлива: слишком велики затраты на их изготовление.
Регуляция транскрипции и трансляции. Клетки различных тканей живых организмов сильно отличаются друг от друга по строению и функциям, но все они произошли от единственной зиготы (оплодотворенной яйцеклетки) в результате многократного деления.
Перед каждым актом деления ДНК реплицируется в две точные копии. Ученым удалось экспериментально доказать, что дифференцированные клетки организма имеют одинаковый набор ДНК, не утрачивая ни одной из его частей в процессе делений.
В 1958 г. Ф. Стюарт впервые вырастил из единственной клетки взрослой моркови целое растение. В 1968 г. Дж. Гордону удалось пересадить ядра из клеток кишечника головастика в яйцеклетки лягушки, лишенные собственных ядер, и вырастить из них нормальных лягушек. Следовательно, клетки всех тканей организма имеют один и тот же набор генов, одну и ту же информацию о строении белков.
Однако клетки каждой ткани производят свои белки, свои ферменты. Гемоглобин образуется только в эритроцитах, белковый гормон роста синтезируется только в клетках гипофиза, зрительный белок опсин — в клетках сетчатки глаза, а инсулин — в клетках поджелудочной железы.
Такое разнообразие происходит по причине того, что клетки каждой ткани реализуют только свою часть информации ДНК. В разных клетках происходит транскрипция разных участков ДНК, синтезируются разные иРНК, по которым воспроизводятся разные белки. Более того, специфичные для клеток белки производятся не все сразу, а по мере необходимости. В организмах имеются удивительно точные механизмы "включения" и "выключения" генов на разных этапах жизненного цикла клетки от ее появления до деления.
Геном эукариот устроен намного сложнее генома прокариот. Это связано с увеличением сложности регуляции генетических процессов, а не с увеличением количества структурных белков и ферментов. В клетках высокоразвитых организмов только 10% всех генов ответственны за синтез ферментов и структурных белков, остальные 90% составляют "административный аппарат" клетки.
Кроме того, существуют системы, регулирующие синтез веществ в организме как едином целом. В клетках желез внутренней секреции вырабатываются гормоны, которые разносятся с кровью по всему телу. Эти гормоны регулируют процессы синтеза иРНК и трансляцию именно в тех клетках, для которых они предназначены. На поверхности мембраны клеток есть рецепторы для "своих" гормонов. Связываясь с рецепторами, гормоны управляют активностью различных систем клетки, регулирующих обмен. В результате может изменяться как транскрипция конкретных генов, так и синтез белков на рибосомах.
Даже синтезированные иРНК могут долгое время не транслироваться, если нет "команды" от гормонов. Каждый гормон через определенные системы клеток активирует свои гены. Так, адреналин дает сигнал на синтез ферментов, расщепляющих гликоген до глюкозы, интенсивно потребляемой мышцами при физической нагрузке. Инсулин выполняет обратную функцию, он участвует в синтезе гликогена из глюкозы в клетках печени.
Как транскрипция, так и трансляция могут подавляться различными химическими веществами, относящимися к классу антибиотиков (греч. anti против + bios жизнь). Например, эритромицин и стрептомицин подавляют синтез белка на рибосомах инфекционных бактерий, а бледная поганка содержит вещество, подавляющее функционирование РНК-полимеразы человека. Прекращение полимеразой синтеза иРНК приводит к тяжелейшим последствиям.
Подведем итоги. В воспроизведении белков в живых существах участвует ДНК, информационная РНК, транспортные РНК для 20 аминокислот (всего 61, по количеству кодонов), рибосомы, состоящие из нескольких рибосомных РНК (3 у прокариот и 4 у эукариот) и десятков различных молекул белка, а также целый комплекс ферментов. Для осуществления синтеза одной белковой молекулы необходим тонкий, специфический подвод энергии посредством более чем 1000 молекул АТФ. Практически вся клетка участвует в синтезе, нарушение строения хотя бы одного из компонентов нарушает процесс воспроизведения белковых молекул.
Для современных ученых удивителен сам факт функционирования этой сложной системы в организме. Доказана абсолютная невозможность самообразования молекул РНК и даже ее более простой составляющей — рибозы. Возможность же самосборки белков в гипотетическом первобытном океане при возникновении в нем жизни квалифицированные биохимики совершенно исключают. Так познание внутриклеточных процессов приводит к мысли о богосотворенности мира и приближает нас к очевидности Божия бытия.
В сельском хозяйстве и садоводстве используют ряд методов искусственного размножения растений. Первые три из описанных ниже методов, а именно — черенкование, прививки и размножение отводками, — традиционные способы, однако на коммерческом уровне они постепенно вытесняются современными методами с применением культуры ткани.
21.3.1. Черенкование
Это несложная процедура, при которой отрезают часть растения и помещают ее в подходящую для роста среду. Со временем черенок пускает корни и вырастает в новое растение. Окоренение можно ускорить гормонами, стимулирующими образование корней. Таким способом обычно размножают широко распространенные комнатные растения — герань и пеларгонию. Другое распространенное комнатное растение — узамбарскую фиалку, или сенполию, размножают отрезанными листьями. Черенкованием размножают черную смородину, отрезая осенью побеги на продажу. Так же размножают хризантемы.
21.3.2. Прививки черенками и почками
Прививка черенками производится путем переноса части растения — привоя — на нижнюю часть другого растения — подвоя. Первоначально прививки делали на яблонях, потому что эти деревья не удавалось выращивать из черенков, а у яблонь, выращенных из семян, наблюдалась слишком сильная изменчивость, так как семена получали путем полового размножения. Теперь эти прививки используют также для размножения других плодовых деревьев, например персиковых, а также слив. Привой выбирают по качеству плодов, а подвой — за такие качества, как устойчивость к болезням и к неблагоприятным условиям.
Розы обычно размножают одним из вариантов этого метода — так называемой окулировкой, при которой в качестве привоя используют не побег, а почку. Новые сорта создают путем полового размножения, однако, как и в случае плодовых деревьев, получить таким способом чистый сорт не удается. Поэтому для сохранения желаемых сортов приходится прибегать к тем или иным способам вегетативного размножения.
21.3.3. Размножение отводками
Отводками размножают растения, выбрасывающие плети, или усы, например землянику. Усы пришпиливают, пока они еще сохраняют связь с родительским растением, а после того, как на них образуются собственные корни, отрезают от него.
21.3.4. Культура ткани, или микрорепродукция

Рис. 21. 11. Культура ткани. А. Растение первоцвета с хорошо развитыми листьями, которые можно разрезать на много мелких частей для клонирования. Б. Фрагменты листа перенесены на агар с соблюдением стерильности. В. Один из фрагментов листа, на котором образовался каллус и новый побег. Г. Побег отделили от фрагмента листа и поместили на достаточно толстый слой агара, чтобы стимулировать рост корней. Д. Молодые клоны пере несли в рыхлую культуральную среду, чтобы усилить развитие корней. Е. Идентичные растения, полученные из одного фрагмента листа путем клонирования.
В начале шестидесятых годов прошлого века было показано, что ядра зрелых растительных клеток содержат всю информацию, необходимую для кодирования целого организма. Проф. Ф. Стьюард из Корнеллского университета (США) показал, что зрелые клетки моркови, помещенные в подходящий культуральный раствор, можно вновь заставить делиться и образовывать новые растения моркови. Эти клетки назвали тотипотентными, поскольку они, даже после достижения зрелости и специализации, сохраняют способность при наличии подходящих условий начать делиться, образуя новые растения.
Преимущества культуры ткани
Ниже перечислены некоторые из важных достоинств метода культуры ткани. Более подробные сведения изложены в конце этого раздела.
1. В ряде случаев можно быстро размножить растения с желаемыми признаками, получая много идентичных копий. Этого нелегко добиться, используя традиционные методы, основанные на половом размножении, особенно если растения адаптированы к перекрестному опылению и аутбридингу (см. далее). Метод культуры ткани имеет важное значение при выращивании ряда зерновых и других сельскохозяйственных растений. Он позволяет быстро размножать новые сорта, полученные путем скрещивания растений.
3. Культивирование клеток не требует много места.
4. Условия культуры ткани исключают возможность заражения растений какими- либо болезнями. Вирусы устраняют описанными ниже методами.
5. Разработаны новые методы создания гибридов путем слияния протопластов, т. е. клеток, освобожденных от своих оболочек. Таким способом удалось получить межвидовые гибриды (например, гибрид картофеля и томата).
6. Эти методы могут оказаться эффективными для получения из растений ценных химических веществ, например лекарств.
Культуральная среда обычно содержит соответствующие питательные вещества и гормоны (см. разд. 16.2). Стандартная среда должна содержать неорганические ионы, необходимые для роста растений, в том числе азот, магний, железо и калий (см. табл. 7.7). Кроме того, нужны сахароза, как источник энергии, и витамины. Эти химические вещества обычно смешивают с агаром, получая желеобразную питательную среду, сходную с той, которую используют для выращивания бактерий и грибов. В культуральной среде должны содержаться незаменимые гормоны — ауксин, стимулирующий рост корней, а также рост клеток в продольном направлении, и цитокинин, стимулирующий рост побегов и клеточное деление. Различные соотношения ауксина и цитокинина оказывают разное влияние на характер развития неспециализированных клеток. Культуру ткани выращивают на поверхности агара в колбах или в чашках Петри.
Температуру, интенсивность и качество света, продолжительность светового дня и влажность регулируют, выращивая культуры в специальных помещениях. Все процедуры проводят в стерильных условиях, так как в культурах могут поселиться бактерии и грибы, растущие быстрее и, следовательно, способные выйти победителями в конкуренции с растениями. Саму растительную ткань стерилизуют, обрабатывая ее поверхность разведенным белильным раствором; остальные материалы также стерилизуют перед употреблением. При работе со всей установкой следует постоянно обеспечивать стерильность, как это делают при микробиологических исследованиях (разд. 12.3).
На рис. 21.12 кратко охарактеризованы основные методы выращивания новых растений с использованием культуры тканей. Фрагменты, взятые от растения, подлежащего размножению, называют эксплантатами. Самый обычный метод состоит в использовании меристематической ткани из верхушечных или пазушных почек. Меристемой называют ткань, в которой клетки все еще продолжают делиться. Другой способ состоит в создании каллуса из немеристематической ткани (рис. 21.13). Каллус — это масса недифференцированных (неспециализированных) клеток. Рост корней или побегов из каллуса или из немеристематической ткани можно стимулировать добавлением ауксинов или цитокининов. На рис. 21.13 видны молодые проростки, возникающие из каллуса. Иногда появляются не побеги и не корни, а зародыши; если поместить их на агаровое желе, они могут дать начало маленьким растеньицам. На рис. 21.11 показаны дальнейшие стадии этого процесса.

Рис. 21.12. Методы получения клонов от одного исходного растения.
Рис. 21.13. Крошечные растения табака, развившиеся из культуры каллусной ткани на стерильном агаре.
Растения, свободные от вирусов
Вирусы могут распространиться по всему растению, и предотвратить их переход с одного растения на другое при размножении традиционными методами очень трудно. Однако обычно вирусы не проникают в верхушечную меристему. Поэтому для получения растений, свободных от вирусов, методом клонирования можно использовать меристемы. Тепловая обработка меристем позволяет повысить уверенность в отсутствии вирусов. Следует создавать фонды меристем, свободных от вирусов, с тем чтобы можно было по мере надобности получать новые растения. Это позволяет снизить расходы по созданию защищенных от инфекций теплиц и служит более надежным способом предотвращения распространения болезней, чем традиционные методы. Так размножают картофель, плодовые деревья, некоторые луковичные и декоративные растения.
Получение безвирусного картофеля
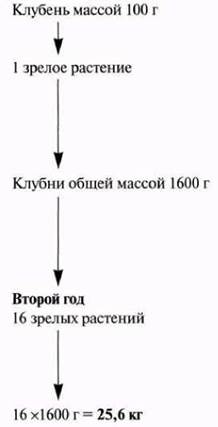
Культуру ткани используют в широких масштабах для разведения декоративных растений, плодовых деревьев и в плантационных хозяйствах, выращивающих такие культуры, как масличная и финиковая пальмы, сахарный тростник или бананы; однако в отношении сельскохозяйственных культур этот метод применяли мало. Единственное исключение составляет картофель (табл. 21.1). Одной из причин этого была возможность получить таким образом растения, свободные от вирусов. Повторные пересевы меристемы позволяют получать большое число маленьких растений, используемых затем для получения мини-клубней, величиной с горошину (рис. 21.14), которые можно высевать подобно семенам. Одно растение способно давать в год свыше полумиллиона мини-клубней. Это позволяет ускорить процесс интродукции новых сортов картофеля, обладающих, например, устойчивостью к серьезной вирусной болезни — скручиванию листьев.
Таблица 21.1. Сравнение традиционных методов размножения клубней картофеля с методом культуры ткани (микроразмножение)

(По данным табл. 12.3 в кн. Molecular Biotechnology, 2nd ed., S. D. Primrose (1991), Blackwell, reproduced from Mantell et al. (1985).)

Рис. 21.14. Мини-клубни картофеля выглядят совершено так же, как обычные клубни, только размеры их не больше размеров горошины.
Картофель считается одним из важнейших культурных растений. Он занимает четвертое место в мире среди сельскохозяйственной продукции после таких злаков, как рис, пшеница и кукуруза. В последнее время его популярность растет в азиатских странах. До недавнего времени можно было интродуцировать новые сорта только путем кроссбридинга растений с желаемыми признаками. Однако существует несколько подвидов картофеля и не все они скрещиваются между собой. Среди тех, которые скрещиваются, некоторые дают только стерильные гибриды. Кроме того, выведение стабильного нового сорта путем повторных генетических скрещиваний занимает обычно 10—15 лет.
Используя культуру ткани, можно получать в массовых количествах как дикорастущие родичи картофеля, так и культурные сорта. Теперь новые сорта картофеля селекционеры способны создавать очень быстро путем переноса полезных генов от, например, дикорастущих родичей культурного сорта или даже от совершенно неродственных растений в отдельные клетки, используя методы генетической инженерии. В наиболее распространенном методе в качестве вектора используют Agrobacterium (см. гл. 25). Затем из трансформированных клеток в культуре ткани можно выращивать небольшие растения и далее размножать их, как было описано выше. Таким образом ген, контролирующий один из белков оболочки вируса, вызывающего скручивание листьев картофеля, был введен в сорта картофеля Дезирэ и Пентленд Скуайр, что эквивалентно вакцинации против этого вируса. Хотя при этом картофель все же может заразиться, однако в таком картофеле вирус размножается гораздо медленнее, чем обычно, и у растения либо наблюдаются незначительные проявления болезни, либо их нет вовсе.
Другой метод, с успехом применяемый к картофелю, состоит в слиянии двух соматических (неполовых) клеток от разных сортов. Для этого у клеток сначала удаляют клеточные стенки, чтобы получить голые протопласты. Образующиеся в результате слияния протопластов соматические гибриды можно выращивать в культуре ткани. Это могут быть гибриды между двумя сортами, которые не удалось бы скрестить половым путем ввиду их несовместимости. Таким образом из дикорастущего картофеля были получены промышленные сорта, устойчивые к вирусу скручивания листьев и к холоду.
Преимущества метода культуры ткани
1. Быстрое размножение. На образующихся при клонировании побегах обычным порядком развиваются почки. Эти почки можно использовать для генерации новых побегов с помощью того же метода культуры ткани. Путем непрерывного повторного клонирования почек число потенциальных растений умножается на каждой стадии. В результате в течение определенного периода от одного побега можно получить тысячи или даже миллионы растений. Это намного быстрее, чем при использовании традиционных методов разведения, так что новые сорта могут быть интродуцированы на несколько лет раньше.
2. Генетическое единообразие. Растения, получаемые при использовании метода культуры ткани, генетически идентичны; иными словами, все они обладают желаемыми признаками исходного растения и размножаются в чистоте. Такие растения (т. е. растения, гомозиготные по желаемым признакам) очень трудно получить при половом размножении.
3. Здоровые растения. Как уже объяснялось выше, используя для размножения меристематические ткани, можно избежать вирусных болезней. Поскольку все процедуры проводятся в стерильных условиях, растения не подвержены опасности заражения поверхностными бактериями и грибами, среди которых могут быть и болезнетворные формы.
4. Культура ткани требует относительно немного места по сравнению с выращиванием растений в теплицах или на полях.
5. Метод культуры ткани не зависит от сезонных изменений погодных условий, т. е. растения с заданными свойствами можно получать в любое время года, тогда, когда их можно продать дороже всего.
6. Возможен более тщательный контроль за развитием растений, что гарантирует единообразие продукта, предлагаемого покупателю.
7. Некоторые растения, например бананы, стерильны и их можно размножать только бесполым путем.
8. Семена некоторых растений, например определенных орхидных, трудно проращивать. Их надежнее размножать бесполым путем.
9. Культура ткани в сочетании с методами генетической инженерии позволяет получать трансгенные растения (см. разд. 25.4).
10. Поскольку культуры ткани не громоздки, их экономично транспортировать по воздуху, что расширяет возможности международной торговли.
Недостатки метода культуры ткани
1. Метод требует больших затрат рабочей силы и менее удобен, чем высеивание семян. Кроме того, он требует достаточно высокой квалификации. Его применение в широких масштабах порождает определенные проблемы в плане организации и обучения персонала. При этом значительно возрастает и стоимость полученного продукта. Обычно применение этого процесса экономически выгодно только для дорогих культур, таких как декоративные растения, и нерентабелен для дешевых культур, например моркови.
2. Необходимость поддерживать стерильные условия. Это еще больше повышает расходы и усложняет весь процесс.
3. Растения, полученные из культур каллуса, иногда претерпевают генетические изменения. Небольшая доля этих изменений может оказаться коммерчески выгодной, но в большинстве случаев они нежелательны.
4. Поскольку клоны генетически идентичны, сельскохозяйственные культуры сильно подвержены заражению новыми болезнями и очень чувствительны к изменениям условий среды. Эти факторы могут полностью уничтожить некоторые культуры.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Под метаболизмом понимают постоянно происходящий в клетках живых организмов обмен веществ и энергии.Энергетический обмен в клетке.Ионы водорода свободно проходят через эти поры, и синтез АТФ в клетках бурого жира не происходит.В процессах метаболизма реализуется наследственная информация.Клетка синтезирует только те вещества, которые записаны в ее генетической программе.Их активность падает, и клетки заменяют их новыми.Матричные процессы составляют основу способности живых организмов к воспроизведению.Синтез небелковых соединений клетка осуществляет в два этапа.В 1958 г. Ф. Стюарт впервые вырастил из единственной клетки взрослой моркови целое растение.Связываясь с рецепторами, гормоны управляют активностью различных систем клетки, регулирующих обмен.
(Если ссылка на скачивание файла не доступна - дайте нам знать об этом в комментариях либо через форму обратной связи)
Читайте также: