
В семязачатке цветка мегаспора прорастает и образует
Добавил пользователь Владимир З. Обновлено: 19.09.2024
Внутри завязи располагаются семязачатки, число которых может варьироваться от одного до нескольких миллионов. Семязачаток – многоклеточное образование семенных растений, из которого развивается семя. Семязачаток состоит из нуцеллуса (мегаспорангия), окруженного одним или двумя интегументами (покровами), которые на верхушке не смыкаются, образуя узкий канал – микропиле. Через этот канал пыльцевая трубка проникает к зародышевому мешку. Основание нуцеллуса, от которого отходят интегументы, называется халазой. Семязачаток прикрепляется к плаценте с помощью фуникулюса (семяножки). Место прикрепления семязачатка к семяножке называют рубчиком.
Семязачаток возникает на плаценте в виде небольшого бугорка. После достижения им определенной величины у его основания возникают интегументы в виде одного-двух кольцевых валиков, а в апексе вычленяются одна или несколько археспориальных клеток. Археспориальная клетка превращается в мегаспороцит. Далее она и делится путем мейоза и образует четыре гаплоидные мегаспоры, составляющие линейную тетраду. Этот процесс носит название мегаспорогенеза. Дальнейшее развитие обычно получает только одна (чаще нижняя) мегаспора, которая дает начало женскому гаметофиту (зародышевому мешку), а три другие мегаспоры дегенерируют.
Выделено 16 типов зародышевых мешков. Наиболее распространен зародышевый мешок Polygonum-типа. При формировании данного типа гаметофита в мегаспоре трижды происходит митоз. Образуется восемь свободных ядер; по три ядра отходят к разным полюсам клетки, окружаются собственной протоплазмой и образуют клетки, покрытые сначала лишь мембраной, а позднее и тонкой оболочкой. Три верхние клетки называют яйцевым аппаратом. Средняя из них становится яйцеклеткой, две другие – синергидами. Три клетки, расположенные на халазальном полюсе, – антиподы. Два ядра, оставшихся в центральной части зародышевого мешка, называют полярными. До или после проникновения пыльцевой трубки в зародышевый мешок они сливаются во вторичное (дип-
Двойное оплодотворение у покрытосеменных растений и его биологический смысл. Отклонения от нормального оплодотворения.
Пыльцевое зерно, попав на рыльце пестика, начинает прорастать. При этом его содержимое выпячивается через поры экзины и образует пыльцевую трубку. Между опылением и оплодотворением проходит различное время. Ядро клетки пыльцевой трубки и два спермия находятся на растущем конце трубки. Достигнув завязи, пыльцевая трубка направляется к семязачатку и проникает в него чаще всего через микропиле (порогамия), в редких случаях – через халазу (халазогамия). Дальше трубка растет по направлению к зародышевому мешку. Оболочка пыльцевой трубки растворяется – и спермии оказываются внутри зародышевого
мешка. Один из них сливается с яйцеклеткой, другой – с диплоидной центральной клеткой. В результате образуются диплоидная зигота и триплоидная клетка, которая дает начало эндосперму. Происходит так называемое двойное оплодотворение – характерный процесс покрытосеменных растений. Из оплодотворенного семязачатка впоследствии формируется семя. Типичный способ оплодотворения называется амфимиксисом.
1. Апомиксис – формирование зародыша из неоплодотворенных элементов зародышевого мешка или семязачатка. Различают следующие его типы:
а) партеногенез – развитие зародыша из неоплодотворенной яйцеклетки с редукционным делением материнской клетки (1n) или без него (2n);
б) апогамия – развитие зародыша (1n или 2n) из антипод или синергид;
в) апоспория – утеря мегаспорогенеза и развитие зародышевого мешка из вегетативной клетки нуцеллуса или интегумента с последующим врастанием внутрь зародышевого мешка.
2. Полиэмбриония (многозародышевость):
а) истинная – развитие в одном зародышевом мешке нескольких зародышей из одной зиготы в результате ее неправильного деления или расщепления ее верхушечной клетки;
б) ложная– образование зародыша либо в результате развития в семязачатке не одной, а нескольких мегаспор (лилия, манжетка), либо в результате развития дополнительных зародышевых мешков из клеток нуцеллуса или интегумента.
Биологическое значение двойного оплодотворения у покрытосеменных растений.
Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных , где довольно мощный гаплоидный эндосперм развивается независимо от процесса оплодотворения, у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов. Увеличение же уровня плоидности эндосперма до 3n, по-видимому, способствует более быстрому росту этой полиплоидной ткани по сравнению с диплоидными тканями спорофита .
Классификация соцветий. Их биологическое значение.
Соцветие – это система видоизмененных побегов, несущих цветки.
По расположению на растении соцветия могут быть терминальными (находятс на верхушках главных побегов), пазушными (на концах обычно укороченных боковых побегов) и интеркалярными (состоят из чередующихся стерильных и фертильных участков).
По наличию и характеру листьев на осях соцветия делят на фрондозные, брактеозные и эбрактеозные. На осях фрондозных соцветий имеются хорошо развитые зеленые прицветники (фуксия, фиалка трехцветная). Брактеозными называют соцветия, в которых прицветники представлены чешуевидными листьями верховой формации – брактеями (ландыш, сирень). У эбрактеозных (голых) соцветий прицветники вообще редуцированы (пастушья сумка).
В зависимости от степени разветвленности выделяют простые и сложные соцветия. У простых на главной оси располагаются одиночные цветки, а у сложных – частные соцветия, т. е. ветвление достигает трех и более порядков.
В зависимости от особенностей функционирования апикальной меристемы выделяют открытые и закрытые соцветия. В открытых (бокоцветных, неопределенных) апикальная меристема не формирует цветки и обладает неограниченным ростом. Цветки возникают на боковых осях и распускаются снизу вверх (ландыш, черемуха). В закрытых соцветиях (верхоцветных, определенных) апикальная меристема расходуется на образование верхушечного цветка (чистотел, барбарис), а цветки распускаются сверху вниз.
В основу общей морфологической классификации соцветий положены два признака: способ ветвления осей и степень их разветвленности. Соответственно выделяют рацемозные, цимозные, составные соцветия и тирсы.
Рацемозные соцветия могут быть простыми (цветки сидят непосредственно на главной оси соцветия) и сложными (цветки сидят на разветвлениях главной оси соцветий). К простым моноподиальным соцветиям относятся следующие:
кисть, когда цветки расположены на удлиненной оси, имеют цветоножки (черемуха);
колос, сходный с кистью, но с сидячими цветками (подорожник);
початок – колос с толстой мясистой осью (кукуруза);
головка, сходная с кистью, но при этом главная ось сильно укорочена, расширена, цветки сидячие или на коротких цветоножках (клевер).
щиток, у которого на укороченной оси развиваются нижние цветки на более длинных цветоножках, верхние – на более коротких, благодаря чему все цветки расположены почти в одной плоскости (спирея);
зонтик, главная ось которого сильно укорочена, цветоножки выходят из ее верхушки и имеют одинаковую длину (проломник);
корзинка, сидячие цветки которой расположены на очень уплощенном и расширенном конце укороченной главной оси (сложноцветные).
К сложным рацемозным соцветиям относятся следующие:
сложный колос, на главной оси которого сидят элементарные колоски (пшеница);
двойная кисть, на главной оси которой сидят пазушные простые кисти (донник);
метелка, отличающаяся от двойной кисти более обильным ветвлением, а также тем, что в нижней части она более ветвистая, чем в верхней (сирень);
сложный зонтик, главная ось соцветия которого укорочена, а от нее отходят несколько осей, несущих на конце простые зонтики (укроп, борщевик).
Цимозные соцветия характеризуются симподиальным ветвлением, относятся к сложным закрытым и представлены следующими типами:
монохазий, т. е. ось первого порядка заканчивается цветком, под которым закладывается одна ось второго порядка. Она перерастает ось первого порядка и также заканчивается цветком, ниже закладывается следующая ось. В зависимости от порядка заложения боковых осей различают завиток и извилину. В завитке все цветки направлены в одну сторону (незабудка). В извилине боковые оси с цветком отходят поочередно в противоположные стороны (гладиолус);
дихазий, т. е. ось первого порядка несет на верхушке цветок, под ним образуются, обычно супротивно, две боковые оси, также заканчивающиеся цветками. Под цветком на боковых осях могут закладываться по две оси третьего порядка и т. д. (герань, гвоздика);
плейохазий, т. е. из каждой оси, несущей верхушечный цветок, выходит более двух ветвей, перерастающих главную ось (белоголовник).
К сложным относятся также составные (агрегатные) соцветия и тирсы.
У составных характер ветвления главной и боковых осей различен. Например, если главная ось ветвится по типу кисти, а боковые частные соцветия представлены корзинками, то такое соцветие называется кистью корзинок. Наиболее распространенными типами составных соцветий являются следующие: щиток корзинок, кисть корзинок, кисть зонтиков, метелка колосков и т. д.
Тирс – это соцветие, имеющее моноподиально ветвящуюся главную ось, несущую боковые цимозные соцветия (монохазии и дихазии).
Биологическая роль соцветий.
Образование соцветий является высокой специализацией к опылению. Как при энтомофилии, так и при анемофилии вероятность опыления цветков в соцветиях возрастает. К тому же цветки в соцветиях распускаются не одновременно, а последовательно, что удлиняет период возможного опыления. В соцветиях повышается гарантия сохранения завязи от повреждения насекомыми и в случае гибели цветков под влиянием неблагоприятных факторов.
Опыление растений. Самоопыление и перекрестное опыление. Приспособления растений к перекрестному опылению.
Опылением называется перенос пыльцы на рыльце пестика.
Различают два основных типа опыления: автогамию, или самоопыление, при котором опыление происходит в пределах одного цветка, и аллогамию (перекрестное опыление) – перенос пыльцы с одного цветка на другой. Если цветки находятся на одном растении, перекрестное опыление называют гейтоногамией, если на разных – ксеногамией. Гейтоногамия генетически эквивалентна автогамии, однако требует тех или иных агентов для переноса пыльцы.
Перекрестное опыление встречается в природе гораздо чаще и обеспечивает генетически более разнообразное потомство. Самоопыление является резервной возможностью опыления в том случае, когда по каким-то причинам (погодные условия, отсутствие опылителя и др.) не произошло перекрестное опыление. В обоеполых цветках есть различные приспособления, препятствующие самоопылению. Обычно таким приспособлением служит дихогамия – разновременность созревания тычинок и пестика (более раннее созревание пыльцы называют протерандрией, рыльца – протерогинией).
У некоторых видов дихогамия сопровождается гетеростилией (разностолбчатостью): у одних цветков тычинки имеют короткие тычиночные нити, а столбик пестика длинный, у других – наоборот. Кроме того, приспособлениями к перекрестному опылению являются самостерильность (неспособность пыльцы прорастать на рыльце пестика того же цветка), раздельнополость цветков, двудомность.
(от мегаспора и . генез), развитие у разноспоровых высших растений мегаспор (или мегаспориальных ядер) в мегаспорангии из мегаспороцита в результате мейоза. У большинства разноспоровых растений мейоз сопровождается последоват. Заложением клеточных стенок, так что после первого деления образуется диада (2 клетки), а после второго — тетрада (4 клетки) обособленных гаплоидных мегаспор. У бессеменных растений каждая мегаспора прорастает в жен. гаметофит; у семенных развивается обычно одна, а три других отмирают. При описанном ходе М. зародышевый мешок развивается из одной мегаспоры и наз. моноспорическим. В др. случаях при М. заложение клеточных перегородок может быть подавлено. Иногда клеточная перегородка закладывается только после первого деления мейоза, а после второго не образуется. В этом случае М. завершается образованием диады, каждая из клеток к-рой содержит по два свободных мегаспориальных ядра. Такая двуядерная клетка соответствует двум необособивщимся мегаспорам и из неё развивается двухспоровый (биспорический) зародышевый мешок (напр., у лука, ландыша, нек-рых амариллисовых). В др. случаях (у тюльпана, лилии, майника) оба деления мейоза не сопровождаются заложением клеточных перегородок и весь мегаспороцит превращается в клетку с 4 свободными ядрами (4-ядерный ценоцит). Такие ценоциты дают начало четырёхспоровым (тетраспорическим) зародышевым мешкам. Моноспорич. зародышевые мешки рассматриваются как исходные и характерны для 80% исследованных покрытосеменных. Двух- и четырёхспоровые зародышевые мешки считаются производными, возникшими позднее в ходе эволюции. (см. ЗАРОДЫШЕВЫЙ МЕШОК).
Мегаспорогенез у резухи кавказской (Агаbis caucasica): 1 — недифференцированный бугорок семяпочки с двумя археспориальными клетками (a); 2 — развивающиеся мегаспороциты (б): 3 — стадия формирования тетрад (второе деление мейоза).
Полезное
Смотреть что такое "МЕГАСПОРОГЕНЕЗ" в других словарях:
мегаспорогенез — мегаспорогенез … Орфографический словарь-справочник
мегаспорогенез — макроспорогенез Развитие у гетероспоровых высших растений мегаспор в мегаспорангиях в результате мейоза; обычно из макроспороцита (или материнской клетки мегаспор) образуется тетрада мегаспор; у низших растений могут образовываться диады (с… … Справочник технического переводчика
мегаспорогенез — ЭМБРИОЛОГИЯ РАСТЕНИЙ МЕГАСПОРОГЕНЕЗ – процесс образования тетрады мегаспор с гаплоидным набором хромосом в каждой мегаспоре … Общая эмбриология: Терминологический словарь
мегаспорогенез — megasporogenezė statusas T sritis augalininkystė apibrėžtis Makrosporų susidarymas. atitikmenys: angl. macrosporogenesis; megasporogenesis rus. макроспорогенез; мегаспорогенез ryšiai: sinonimas – makrosporogenezė … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
мегаспорогенез — процесс образования мегаспор в мегаспорангиях разноспоровых растений … Анатомия и морфология растений
МЕГАСПОРОГЕНЕЗ — см. макроспорогенез … Словарь ботанических терминов
Макроспорогенез мегаспорогенез — Макроспорогенез, мегаспорогенез * макраспорагенез, мегаспорагенез * macrosporogenesis or megasporogenesis процесс образования макроспор (см.) … Генетика. Энциклопедический словарь
Спорообразование — спорогенез, процесс образования спор (См. Споры). У растительных организмов прокариотов (См. Прокариоты), клетки которых не имеют типичных ядер, споры могут возникать: из целой клетки, накопившей питательные вещества и утолщившей оболочку … Большая советская энциклопедия
Женский гаметофит (зародышевый мешок) — Женский гаметофит цветковых растений обычно называется зародышевым мешком. Возникновение этого названия восходит к тем временам, когда еще не была ясна морфологическая природа чрезвычайно своеобразного женского гамотофита цветковых… … Биологическая энциклопедия
Строение и типы семязачатков. Семязачаток состоит из нуцеллюса ( ядра), и ингументов (покровов) , края которых образуют узкий канал ( микропиле), через который пыльцевая трубка проникает к зародышевому мешку, а также фуникулюса (семяножки), посредством которого семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где нуцеллюс и интегументы сливаются, называют халазой.
Различают 5 основных типов семязачатков:
п р я м о й семязачаток. Микропиле находится на одной оси с рубчиком фуникулюсом. Семяножка очень короткая ( гречишные, ароидные).
о б р а щ е н н ы й семязачаток. Он повернут на 180 градусов, так что микропиле и рубчик рассоложены рядом, причем микропиле обращено к плаценте. Это самый распространенный тип семязачатков среди цветковых
п о л у п о в е р н у т ы й семязачаток. Нуцеллюс с интегументами повернуты на 90 градусов ( примуловые, норичниковые).
О д н о с т о р о н н е и з о г н у т ы й семязачаток. Характеризуется односторонним разрастанием нуцеллюса и интегументов. Встречается у бобовых и центросеменных.
Д в у с т о р о н н е и з о г н у т ы й семязачаток . Нуцеллюс имеет подковообразную форму. Встречается у отдельных представителей тех семейств, где обычно односторонне изогнутые семязачатки.
Количество семязачатков в завязи варьирует от одного до огромного количества.
В завязи Gymbidium из семейства орхидных обнаружено до 2 000 000 семязачатков.
Мегаспорогенез и мегагаметогенез осуществляется в семязачатках. Снаружи семязачаток покрыт ингументами, которые не смыкаются, оставляя отверстие - микропиле или пыльцевход. Конец семязачатка, противоположный микропиле, называется халазой. Под ингументами семязачатка расположен многоклеточный нуцеллюс, или ядро семязачатка. Вскоре после заложения семязачатка и развития нуцеллуса одна из его клеток начинает разрастаться - материнская клетка. Материнская диплоидная клетка приступает к мейозу. В результате мейоза образуется четыре гаплодных клетки. В дальнейшем три мегаспоры отмирают и только одна, обращенная к халазе, остается. Мегаспора всегда одноядерна и гаплоидна. Мегаспора сильно разрастается, ее ядро претерпевает три последовательных митотических деления. После первого деления образуются два ядра, которые перемещаются к двум противоположным полюсам прорастающей мегаспоры - зародышевого мешка, где осуществляется два последующих деления. В результате на каждом из полюсов образуется по четыре ядра. Все восемь ядер гаплоидны. Три ядра у каждого полюса обособляются в клетки. На микропилярном полюсе одна из клеток становится яйцеклеткой, две другие синергидами. В совокупности они формируют яйцевой аппарат. Три клетки на халазальном полюсе называются антиподами. Два оставшихся ядра перемещаются к центру и сливаются.
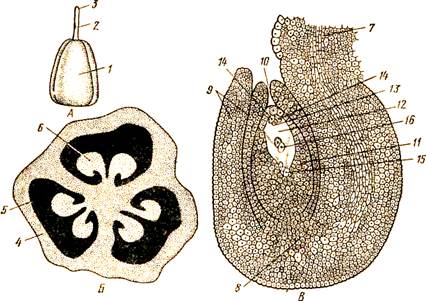
Рис. 115. Гинецей пролески. А — общий вид; Б — поперечный разрез завязи; В — продольный срез семязачатка:
1— завязь, 2 — столбик, 3 — рыльце, 4 — стенка завязи, 5 — гнезда завязи,
6 — семязачаток, 7 — семяножка, 8 — халаза, 9 — интегументы, 10 — микропиле,
Л — нуцеллус, 12 — зародышевый мешок, 13 — яйцеклетка, 14 — синергиды, 15 — антиподы, 16 — вторичное ядро
Так формируется вторичное ядро зародышевого мешка. После этого все компоненты зародышевого мешка готовы к оплодотворению.
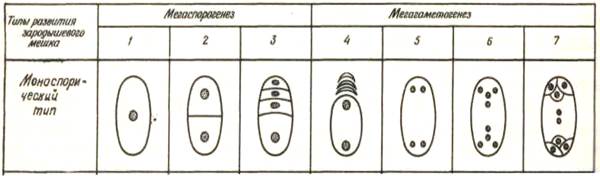 |
Рис.116 Мегаспорогенез и метагаметогенез у покрытосеменных ( схема)
1-материнская клетка спор, 2-6 основные стадии развития зародышевого мешка, 7- зрелый зародышевый мешок.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).


Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Семязачаток или семяпочка, многоклеточное образование у семенных растений, из которого развивается семя. Основные части С. - нуцеллус, интегумент (или интегументы) и семяножка. Нуцеллус возникает в типичных случаях в виде бугорка из клеток плаценты мегаспорофилла (плодолистика). Интегумент закладывается в виде кольцевого валика в основании нуцеллуса и обрастает развивающийся нуцеллус, оставляя над его вершиной узкий канал - микропиле, или пыльцевход, под которым у большинства голосеменных находится пыльцевая камера. Семяножка (фуникулус) соединяет. с плацентой. базальную часть, от которой отходит семяножка, называемая халазой.
В завязи пестика располагаются небольшие образования - семязачатки. Их число колеблется от одного (пшеница, слива) до нескольких миллионов (у архидных). Функции семязачатка - мегаспорогенез (создание мегаспор) и мегагаметогенез (формирование женского гаметофита, процесс оплодотворения). Оплодотворенный семязачаток развивается в семя. Плацента - место прикрепления семязачатка к плодолистику.
нуцеллус, ядро семязачатка;
фуникулюс, семяножка, с помощью которой семязачаток крепится к плаценте;
интегументы, покровы семязачатка, образующие на верхушке нуцеллуса канал;
халаза, базальная часть семязачатка, где сливаются нуцеллус и интегументы;
рубчик - место присоединения семязачатка к семяножке.

Рис.4 Строение семязачатка
ортотропный - прямой, фуникулюс и микропиле расположены на противоположных концах оси семязачатка (гречишные, ореховые);
анатропный (обратный) - нуцеллус повернут по отношению к прямой оси семязачатка на 180, вследствие чего микропиле и фуникулюс расположены рядом (Покрытосеменные)
гемитропный (полуповернутый) - семязачаток повернут на 90, вследствие чего микропиле и нуцеллус расположены по отошению к фуникулюсу под углом 90 (первоцветные, норичные).
кампилотропный (односторонне изогнутый) - нуцеллус изогнут односторонне микропилярным концом, соответсвенно микропиле и фуникулюс расположены рядом (бобовые, мальвовые)
амфитропный (двусторонне изогнутый) - нуцеллус изогнут двусторонне в виде подковы, при этом микропиле и фуникулюс располагаются рядом (тутовые, ладанниковые).

Рис.5 Основные типы семязачатков
Развитие семязачатка
Мегаспорогенез протекает в женской репродуктивной сфере - в гинецее. Морфологически гинецей представлен пестиком (или пестиками). В состав пестика входят: рыльце, столбик и завязь. Внутри завязи содержатся семязачатки (один или несколько). Внутреннее содержимое семязачатка представляет собой нуцеллус. Покровы семязачатка образованы двойным или одиночным интегументом. В нуцеллусе семязачатка имеется одна археспориальная клетка (2n), способная делиться путем мейоза (у ив и некоторых других растений археспорий многоклеточный). В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре гаплоидные мегаспоры (n). Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит). Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра - одно центральное диплоидное ядро; два ядра - две клетки-синергиды; одно ядро становится ядром яйцеклетки.
Когда к семязачатку приближается пыльцевая трубка, он это заранее "чувствует" и готовится ее встретить. Клетки-спутницы начинают выделять слизистые вещества. Тем временем пыльцевая трубка растет, преодолевая сопротивление клеток стенок завязи. Наконец, она достигает микропиле. Происходит "драматический" процесс: пыльцевая трубка протыкает (и при этом убивает) одну из клеток-спутниц. Оба спермия покидают пыльцевую трубку. Участь вегетативной клетки пыльцевого зерна незавидна, она вскоре погибнет. Наблюдать этот процесс очень трудно, но еще труднее понять, что происходит при оплодотворении.
Каждый раз, когда пыльцевая трубка наблюдалась в соприкосновении с зародышевым мешком, оба мужские половые ядра также наблюдались в содержимом зародышевого мешка. Мужские ядра лежат сначала близко друг от друга.
Мужские ядра затем отделяются друг от друга, причем одно проникает к яйцеклетке, а другое тесно прикладывается к одному из еще не слившихся в это время полярных ядер, именно к сестринскому ядру яйцеклетки.
В то время, как мужское ядро все более и более тесно прилегает к ядру яйца, полярное ядро, копулирующее с другим мужским ядром, направляется навстречу к другому полярному ядру, с которым встречается в середине зародышевого мешка.
Только после прохождения профазы деления ядра сливаются. "
Чтобы объяснение было понятнее, Сергей Гаврилович сделал к рисунки, очень понравившиеся столичным ботаникам. Они отвезли их, чтобы показать на заседании Петербургской Академии.
Почему же этому выступлению ученые придали такое большое значение? Считалось, что оплодотворение у растений происходит точно так же как у животных. Один спермий и одна яйцеклетка должны давать зиготу, из которой развивается новое растение. Не то чтобы ботаники не видели двух спермиев, движущихся в пыльцевой трубке к зародышевому мешку. Считалось, что это - какая-то ненормальность, "типичное" растение должно иметь не два, а один спермий, "типичное" оплодотворение должно быть одинаковым и у растении и у животных. Оказалось, что эти представления были неправильными. Оплодотворение цветковых растений совершенно не похоже на оплодотворение не только животных, но и других растений. Процесс оплодотворения с помощью двух спермиев получил название двойного оплодотворения. Сразу же после открытия Навашина ученые бросились к своим препаратам. Оказалось, что многие уже видели двойное оплодотворение, но считали его уродливым процессом, якобы не дающим нормальных семян. Ученые стали присылать Сергею Гавриловичу поздравления. А один ученый даже подарил свои старые препараты, на которых было хорошо видно двойное оплодотворение.
Итак, С.Г. Навашин открыл удивительное явление. Зачем происходит оплодотворение яйцеклетки было понятно: чтобы получилась зигота, а из нее - новое растение. Но зачем же оплодотворять центральную клетку зародышевого мешка? Оказалось что именно из этой клетки развивается питательная ткань семени - эндосперм. Навашин предположил, что без мужского ядра эндосперм образоваться не может. Как же проверить это предположение? Ведь из пыльцевой трудки невозможно вытащить один из спермиев, не нарушая процесс оплодотворения.
Сергей Гаврилович решил исследовать растения, у которых нет эндосперма. Вдруг у них нарушено оплодотворение центральной клетки? Такие растения отыскались в семействе Орхидных (Orchidaceae). Семена орхидных очень мелкие, и даже прорости самостоятельно не могут (а прорастают только при помощи грибов базидиомицетов, образуя микоризу). В пыльцевой трубке орхидных Навашин увидел два спермия. Один из них оплодотворял яйцеклетку, а второй "пробовал" оплодотворить центральную клетку с двумя полярными ядрами. Но в центральной клетке ядра не сливались друг с другом! Процесс оплодотворения был нарушен и, естественно, эндосперм не мог образоваться. В дальнейших работах ученому удалось показать, что у подсолнечника (Helianthus annuus) и некоторых других растений оплодотворение - двойное. После того, как Навашин нашел двойное оплодотворение у самых разных растений, он сделал вывод, что двойное оплодотворение свойственно всем цветковым растениям.
Взглянем на строение семязачатка после двойного оплодотворения. Клетки обоих его покровов как имели два набора хромосом, так их и сохранили. Оба набора в этих клетках принадлежат материнскому растению. Нуцеллус также несет два материнских наборов хромосом. В зародышевом мешке синергиды погибли, а у антиподов по одному набору хромосом. Эти наборы - тоже материнские. Зигота, образовавшаяся слиянием яйцеклетки и спермия, несет два набора хромосом: один от отцовского, а другой от материнского растения. Самое интересное, что центральная клетка имеет три набора хромосом: один от отцовского растения, и два - от материнского.
После двойного оплодотворения начинается несколько процессов: первичное ядро эндосперма делится, образуя эндосперм, зигота развивается в зародыш, интегументы превращаются в семенную кожуру, а стенка завязи и связанные с ней структуры формируют плод. На ранних стадиях развития последовательность клеточных делений у зародышей двудольных и однодольных сходна; в обоих случаях формируются сферические тела. Позже появляется различие: зародыш двудольных имеет две семядоли, а однодольных - только одну. В семенах некоторых групп покрытосеменных нуцеллус развивается в запасную ткань, называемую перистермом. Некоторые семена содержат и эндосперм и перистерм, так у свеклы (Belta). Однако у многих двудольных и некоторых однодольных вся запасная ткань или большая ее часть поглощается развивающимся зародышем еще до того, как семя впадет в состояние покоя (у гороха, у бобов и др.). Зародыши в таких семенах обычно развивают заполненные питательными веществами мясистые семядоли. Основные питательные вещества, запасаемые в семенах, - углеводы, белки, липиды. Семена голосеменных и покрытосеменных различаются происхождением этих веществ. У первых их дает женский гаметофит, у вторых - эндосперм, не являющийся ни гаметофитной, ни спорофитной тканью.
Развитие семяпочки в семя сопровождается превращением завязи (а иногда и других частей растения) в плод. В ходе этого стенка завязи (околоплодник, или перикарарпий) часто утолщается и дифференцируется на отдельные слои - внешний экзокарпий (внеплодник), средний мезокарпий (межплодник) и внутренний эндокарпий (внутреплодник), - обычно лучше заметные в мясистых, чем в сухих, плодах.
Так происходит развитие гаметофита, оплодотворение и формирование семени у "типичного" цветкового растения, однако мир растений очень разнообразен и у множества покрытосеменных развитие проходит иначе, однако не менее интересно и удивительно. Все эти способы развития помогают каждому растению поддерживать свою стратегию в борьбе за существование; преимущество двойного оплодотворения не ограничивается тем, что растения формируют питательные запасы в семязачатке только при условии оплодотворения (не "растрачивая" силы на формирование питательных запасов в семязачатке, который никогда не будет оплодотворен, как это часто происходит у голосеменных), эти различия в развитии, являющиеся следствием приспособленности растений к окружающим их условиям, и создают то удивительное многообразие форм морфологического и физиологического строения, которое мы и наблюдаем у цветковых.
Читайте также: