
Выведение тетраплоидного сорта ржи какая изменчивость
Добавил пользователь Евгений Кузнецов Обновлено: 19.09.2024
Селекция растений сегодня оперирует обширным набором средств для выведения сортов с улучшенными характеристиками. Ученым при выборе методов селекции нужно учитывать некоторые особенности биологии растений.
1. Большая плодовитость, способность давать многочисленное потомство.
2. Значительный выбор самоопыляемых видов (в частности, у пшеницы, гороха, ячменя и пр.).
3. Возможность размножения с помощью вегетативных органов.
4. Относительная простота искусственного получения форм-мутантов.
Ключевые методы селекции растений
1. Искусственный отбор (который, в свою очередь, делится на индивидуальный и массовый).
2. Гибридизация (инбридинг, аутбридинг, межлинейная гибридизация).
5. Искусственный мутагенез.
6. Использование соматических мутаций.
7. Методы клеточной инженерии (или соматическая гибридизация).
8. Методы генной инженерии.
Очень специфическими методами селекции растений являются методы получения соматических мутаций, а также соматических гибридов. Большим потенциалом в селекции растений обладает межлинейная гибридизация.
Искусственный отбор и гибридизация
1. Главные методы, применяемые в селекции растений, — искусственный отбор и гибридизация.
3. Гибридизация резко улучшает качества и разнообразие того природного материала, с которым экспериментирует селекционер.
4. Однако в большинстве случаев метод гибридизации сам по себе не приводит к целенаправленному изменению признаков. Иначе говоря, скрещивания без искусственного отбора дадут небольшой эффект.
5. В селекции растений используется преимущественно массовый искусственный отбор. Реже может использоваться индивидуальный отбор.
6. Применяются два вида гибридизации. Инбридинг проводят до получения гетерозисных гибридов, для закрепления наследственных свойств внутри одной чистой линии.
7. При аутбридинге у растений возникают бесплодные гибриды, требующие полиплоидии для восстановления плодовитости.
1. Гетерозис часто применяется селекционерами для повышения урожайности растений. Он обеспечивается так называемой межлинейной гибридизацией чистых линий.
Полиплоидия
1. Полиплоидия очень распространена в селекции растений.
2. Полиплоидные особи демонстрируют более высокую жизнеспособность, чем диплоидные.
3. Кроме того, избыток хромосом, полученных после полиплоидии, повышает устойчивость растений к бактериям, вирусам, грибам и прочим патогенным организмам, а также к различным неблагоприятным факторам (химическому воздействию, радиации и пр.). Когда повреждаются гомологичные хромосомы, одна или даже две, аналогичные повреждениям не подвергаются.
5. В 1930-е годы селекционер Николай Васильевич Цицин скрестил пшеницу и пырей, получив высокоурожайные и устойчивые к полеганию пшенично-пырейные гибриды.
Искусственный мутагенез
1. Метод начал применяться после открытия результатов воздействия на растения различных мутагенов — химических факторов и излучений. Мутагены дают возможность добиваться широкого спектра мутаций.
2. Искусственный мутагенез подталкивает геномные мутации, которые приводят к полиплоидии. В частности, полиплоидные формы получают, обрабатывая алкалоидом колхицином семена во время прорастания. Если обработать зародыш во время первого митоза его клеток (на стадии метафазы), микротрубочки разрушатся, хромосомы разойдутся случайно. Одна из клеток может стать не диплоидной, а тетраплоидной. Вторая вовсе не получит хромосом. Тетраплоид далее будет делиться митозом.
Отдаленная гибридизация
1. Аутбридинг, или же отдаленная гибридизация — метод скрещивания неродственных растений, которые относятся к разным видам. Неродственным гибридам свойственна стерильность, поскольку у них нарушается ход мейоза.

( местная репродукция Белта х Тетра к ороткая ) х Шатиловская тетра .
Метод семейно - группового отбора сочетает преимущес тва ин -
Рис . 9.10. Схема се мейно - групповог о отбора : 1 – исходный материал ;
2 – группа сходны х семей , высеянная на изолированном у частке ;
3 – испытание лу чших семей ; 4 – размноже ние потомства лу чших семей ;
5 – государствен ное сортоиспытани е ; забракованные деля нки перечёркнуты
Семейно - групповой отбор – вид индивидуального отбор а у пере -
крёстноопылит елей , п ри котором родоначальные элитные растения
разбиваются на несколько групп по о тноси тельно схо дным х озяй -
ственно - биологическим признакам . Каждая группа семей высевается
на изолированной площадке . Переопыление потом ств растений про -
исходит в пределах группы . При использовании семейно - группового
отбора выравненность потомств д остигается медленно , но устраняет -
ся негативный э ффект б лизкородственного разведения . Примером
использовани я данного метода в селекции является сорт диплоидной
гречихи Наташа (200 4), который был выведен многократным семей -
но - групповым от бором из гибридной популяции , полученной при
свободном переопы лении сорта Ирменка и скороспелых крупноплод -
Сорт гречихи Саулык (1998) выведен сочетанием индивидуально -
семейного и семейно - группового отборов из сложной г ибридной по -
пуляции , сформированн ой из крупноплодны х термо стойких ф орм .
В отличие от самоопыляющихся растений селекция перекрёстно -
опыляющихс я видов не может стро иться н а выделении гомозиготных
щихся видов вследствие а утбридинга растения любого сорта имеют
гетерозиготные геноти пы , отличающиеся от других генотипов данной
популяции . Совокупности р екомбина ций генов в общих чертах о ста -
ются сходными из поко лен ия в поколение ; 2 ) принудительное само -
опыление перекрёстноопыляющихся культур приво дит к инцухт -
депрессии потомства . Чтобы избежать инцухт - депрессии , возникаю -
щей при многократн ом индивидуальном отборе , применяют методы
индивидуал ьно - семейного и семейно - группового отборов .
При проведении отбора у перекрёстноопыляющи хся культур об -
ращают внимание не на индивидуальные генотипы растений , а на ге -
нотипы их гамет , их способность обеспечивать наиболее благоприят -
ные комбинации с гаметами друг их растений . У аллогамных видов
при улучшении отдельных количественных признаков ( урожайность ,
размер семени , содержание определённых веществ ) успешно приме -
няют пар ные ск рещивания . В к ачест ве родительской формы в ыби -
рают генетически не родственные особи , п роисходящие из разны х
сортов - популяц ий , с интенсивным проявлением соответствующего
Когда можно полу чить клоновое потомство ( у свёклы , кормовых
трав ), иногда прибегают к топкроссам : одну из р одител ьских форм
скрещивают со всеми остальными , выполняющими р оль материнских
Когда оценку хозяйственной ценности р астений можно сделать
до ц ветения ( у с вёклы , турнепса , моркови и др .), сразу же выбрако -
вывают худшие и средние потомства . Д ля дальнейш его самопроиз -
вольного взаимного с крещивания оставляют то лько самые продук -
Новые пары создают путём отбор а подходящих растений из са -
мых лучших отлич ающихся др уг от друга по происхождению
После нескольких циклов парного скрещивания семена элитных
растений от скрещивания самых лучших пар , выравненных по фено -
типу , с высокой урожайност ью смешивают и высевают совместно .
Таким образом получают новую , улучшенную популяцию .
Если полную характеристику хо зяйственной ц енности можно
дать то лько по сле цветения ( у ржи , гречихи и др . ), необходимо про -
вести большое число парны х скрещиваний , а ценность отдельных пар
испытывать с помощью метода половинок , или резервов .
При проведении возвратных ск рещиваний у пер екрёстноопы -
ляющихся культур во избежание существенного изменения геноти -
пического состава и снижения общей ценности рекуррентного сор та
необходимо повысить масштабы возвратных скрещиваний ( не менее
100–200 р азных особей рекуррентного родителя ). В некоторых случа -
ях аутостерильность или сильная инцухт - депрессия затрудняют по -
ей о т родителя - донора признаком . Поэтому возвратные скрещивания
9.11. Ме т оды о тбора , основа нные на использов ании эффекта
Материнские и отцовские растения в популяциях п ерекрёстни ков
неодинаковы по генотипу , поэтому все особи показывают высокую
степень гетерозиготности . Следовательно , на выражение признаков
могут влиять кроме аддитивны х и неаддити вные эффекты ( домини -
рование и сверхдоминирование ). Так как неаддитивные эффекты свя -
заны с гетерозиготным состоянием , то ва жно применять методы от -
бора , обеспечиваю щие сохранение у отобранных растений высокой
Индивидуал ьная продуктивность о собей в популя циях пере -
крёстников зависит от комбинационной способности их родительских
форм . Комбинационная способность состоит из д вух ко мпонент ов :
ОКС , обусловленной аддитивным действием гено в , и СКС , завися -
Методы отбора при р аботе с аллогамными видами различаются
тем , какая форма комбинаци онной способности у них преобладает .
На и спользовании э ффекта ОКС основаны повторяющийся о тбор на
ОКС и поликросс - тест . В данном случае неаддитивная варианса мно -

го меньше ½ аддитивной ге нетической вариансы . Если неаддитивная
варианса , обусловленная доминированием , больше ½ аддитивной г е -
нетической варианс ы , следует ожидать больший эффект от использо -
вания СКС . Этот эффект применяют в с елекции на гетерозис . Возмо -
жен и одновременный отбор на эффекты ОКС и СКС при и спользо -
Рекуррентный ( повторяющийся ) отбор на ОКС отличается от
повторяющ егося отбора по фенотип у тем , что не является методом
массового отбора , а основан на принципе индивидуального отбора
Сх е ма р ек у рре нт н ог о от б ор а н а об щу ю ко мб ин а ци он ну ю с п ос о
Элитные растения отбираются на о сновани и не только собствен -
ной продуктивности , но и их ОКС с сортом - тестером . Элитные рас -
тения отбирают из и сходного материал а , как при индивидуальном
отборе : сначала по фенотипическому проявлен ию интересующих
признаков . Часть соцветий каждого элит ного растени я изол ируют и
таким образом обеспечи вают принудительное самоопыление . В семе -
нах , полученных путём самоопыления , сохраняется набор генов
элитных растений . Другую часть соцвети й каждого элитного расте -
ния опыляют пыльцой сорта - тестера ( свободноопыляющи йся сорт с
относительно широкой приспособл енностью ). Семена , полученные от
элитных растений , опылённых пыльцой сорта те стера , служат для
изучения ОКС э литных растений . Число о тбираемых элитных р асте -

ний определяется долей ценных растений в популяции и числом ра -
бочих рук . Если затраты труда больше , то обходятся без скрещивания
с тестером , а комбинационную сп особность элитных растений опре -
деляют при свободном опыл ении с другими растениями популяции
исходного материала . Критерием ОКС служит урожайность или дру -
гой количественный признак потомства элитных растений .
Семена элитных растений , полученные от принудительного са -
моопылени я и показавшие наилучшую комбин ационную способ -
ность , смешивают в новую свободнопереопыляющуюся популяцию .
Полученная о т свободного переопыления популяция - синтетик 1 слу -
жит исходным материалом для сл едующ его цикла рекуррентного от -
бора , если синте тик 1 превзошёл по урожайности исходный материал .
Обычно отбор продолжают в течение не скольких циклов . После
2–3 ци клов возможн о п ередать производству улучшенную поп уля -
цию ( сорт ) и в то же время продолж ать дальнейшее улучшение .
Реципрокн ый повторяющийся ( рекуррен тный ) отбор должен
служить для одновременного улучшения ОКС и С КС ( рис . 9.12 ).
Этот способ ис пользуе тся при на личии аддитивной и неаддитив -
ной варианс . Причём степень доминирования главного признака от -
бора должна быть больше единицы . Реципрокный повторяющ ийся
отбор сходен с рекуррентным отбором на ОКС . Разница состоит в
том , что отбор одновременн о происходит в двух популяциях , которые
служат др уг для друга сортом - тестером . В предварительном диал -

лельном скре щивани и некоторых популяций определяют , какие из
них при гибридизации друг с другом имеют наилучшую комбинаци -
онную способность . С н ими должна быть проведена работа методом
Процедура реципрокного повторяю щегося отбора состоит в том ,
что в двух неродственных популяциях , обозначаемых как А и В , о т -
бирают элитные растения и подвергают часть их соцветий принуди -
тельному самоопыл ению . Другую часть – пыльцой сорта - тестера ( со -
Полученные при скрещивании семена элитных растений испыты -
вают на комбинационную способность в полевом о пыте . Для элитных
растений каждой популяции закладывают отдельный опыт . Затем в
каждой популяции проводят отбор растений с луч шей комбинацион -
П олу чен ны е от сам о оп ыл е ни я сем ена эт и х рас те н ий см еш и ва ют и
п олу чаю т дв е ул уч ш ен ны е п опу ляц ии . Ка ж да я по пу ля ц ия п од ве р га ет ся
с во бо дн ом у пе ре оп ы лен ию дл я у ст р ан ен и я ин цу хт - д е пр ес си и и об ес -
п еч ен ия р ек ом би на ц ии ге н оти пов . С е ме на от с во б одн о го пе р ео пы ле ни я
с лу жа т и сх од ны м м ат е ри ал ом дл я сл е дую щ е го ц и кла от б ор а .
Поликросс - тест применяют для отбора на ОКС ( рис . 9.13).
С помощью этого метода можно выделить генотипы , позволяю -
щие образовать прод укти вный с интетический сорт . Поликросс - тест
используют , если , например , по причине самостерильности рек ур -
рентный отбор на ОКС не пригоден или слишком сложен . Применя -
личные зла ковые тра вы ). Из гетерогенного исходного материала с
помощью строгого отбора выдел яют и клонируют соответствующие
цели селекции особи . Клоны образуют для сохранения генотипа
элитного растения и испытания на генотипическую ценность . Для
определения ОКС закладывают п оликроссны е поля . Каждый клон из
наилучших элит ных растений опыляется смес ью пыльцы всех
Каждый к лон в ысаживается в много кратной повторност и ( не ме -
нее 10), распределяются отдельные повторности ка ждого клона рен -
домизированно , так чтобы каждый клон один раз был бы р аспол ожен
по соседству с каждым другим . Высаженные клоны должны цвести
примерно одновременно . Если клоны не полностью самостерильны ,
необходима примерно одинаковая сте пень самостерильности . Д ля
удовлетворен ия этим требованиям нужно от каждого клона получать
больше растений , чтобы каждый клон был предствле н гр уппой н е
Схема закладки поликроссного поля соответствует с хеме блочно -
го о пыт а , причём каждый к лон встречается в каждом блоке один р аз
Полученные при взаимном опылении всех клонов семена соби -
рают по клонам и испытывают на урожайность в по левом о пыте в те -
чение нескольких лет в разных зо нах . Клоны , котор ые при свободном
опылении со всеми другими клонами дают генера тивное по томство
с наивысшей урожайностью , объединяют в синте тический сорт . По
некоторым рекомендациям , самое экономичное число клонов для
При работе с вегетативно размножающими ся культурами ( кар -
тофель , банан , м ногие цветочные , все плодовые и ягодные культуры ,
лесные породы ) любое отобранное ценное родоначальное р астение
независимо от того , гомозиготное оно или гетероз иготное , может

быть широко размножено вегетативным путём и выпущ ено в каче -
стве нового сорта . Это позволяет на лю бом этапе работы выделить
ценные в се лекционном отношении элитные р астения и п утём вегета -
тивного размножен ия закрепить их свойства ( рис . 9.14).
Рис . 9.14. Схема сел екционной работы с вегетативно размножающим ися
Клоном называют вегетативное потомство , полученное от от -
дельного растения или его части . Все особи одного клона генотипи -
чески одина ковы , если внутри него не возникли мутации .
Клоновый отбор – это индивидуальный отбор у вегетативно раз -
множающи хся растений . Как и другие методы отб ора , вк лю чает два
этапа : 1) отбор эл итных растений ; 2) испытание вегетати вного потом -
Проверка результатов отбора ( испыт ание к лонов ) не отличается
от проверки генеративного потомства при индивидуал ьном отборе .
зависимости от типа генетической изменчи вости , и спользуе мой для
1. В естест венной популяции вегетативн о размножающегося в ида
встречаетс я ясно выраженная генетическая из менчивость . В этом
случае отбирают элитные растен ия и вегетатив но размножают каждое
из них , а затем последовательно испытывают их потомства , т . е . кло -
ны . Испытание клонов прово дят так же , как и испытание генератив -
ных потомств , только вместо генеративного размножения происходит
2. Есл и вегета тив но разм нож ающ аяс я в прои звод стве кул ьту ра об -
лад ает способ ност ью раз мно жатьс я и ген ерат ивно , ген оти пич ескую
из мен чиво ст ь можн о резко повы си ть пров еден ие м скре щи ван ия ( с во -
бод ног о пе ре опыл ени я ра зли чны х кл оно в ) и полу чени ем гене рат ивн о -
го пото мст ва . У ал лог амны х вид ов , вс лед стви е г ет еро зиго тн ости кло -
но в , уж е п рост ое са моо пыл ени е при вод ит к расщ еп лени ю , кото рое
уси ли вае тся при сво бодн ом оп ылен ии меж ду разн ыми кло нам и .
3. В клонах могут возникнуть мутации – спонтанно или искус -
ственным путём . В результате клоновые растения ок азываю тся хи ме -
рами , т . е . состоят из генетичес ки разных кл еток и тканей .
1) стимулируют развитие всех поч ек каждой особи . Это до стига -
ется , например , о трезанием верхушки с тебля , что в ызывае т прораста -
Методы селекции растений
1. Какая разница с генетической точки зрения между самоопылением и перекрестным опылением?
2. Что такое полиплоидия?
3. Почему большинство культурных растений размножают вегетативно?
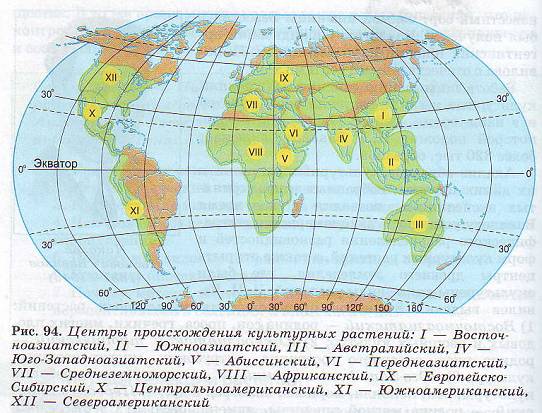
Центры происхождения культурных растений.
Основой успеха селекционной работы в значительной степени является генетическое разнообразие исходного материала. В своей работе селекционеры стараются использовать все многообразие диких и культурных растений.
На необходимость использовать в селекции растений все видовое многообразие флоры нашей планеты указывал еще академик Николай Иванович Вавилов, выдающийся генетик и селекционер. Под его руководством были организованы научные экспедиции в разные регионы Земли для сбора образцов культурных растений, их диких предков и сородичей. В ходе экспедиций было собрано более 160 тыс. образцов разных видов и сортов растений.
В настоящее время эта уникальная коллекция хранится во Всесоюзном институте растениеводства и используется селекционерами в их практической работе. Так, известный сорт озимой пшеницы Безостая-1 был получен в результате гибридизации аргентинских пшениц из коллекции Н. И. Вавилова с отечественными сортами.

Работа по созданию семенных коллекций культурных и диких растений продолжается и в наше время. Сейчас коллекция, начало которой положил Н. И. Вавилов, включает более 320 тыс. образцов.
Анализ образцов культурных растений и их диких предков, собранных в предпринятых экспедициях, позволил в свое время Вавилову установить закономерности географического распределения разновидностей и форм культурных растений, а также открыть центры древнего земледелия, где были окультурены дикие виды растений. Н. И. Вавилов выделил 8 центров происхождения культурных растений: 1) Восточноазиатский — родина сои, проса, гречихи, многих плодовых и овощных культур; 2) Южноазиатский тропический — родина риса, сахарного тростника, цитрусовых, многих овощных культур; 3) Юго-Западноазиатский — пшеница, рожь, бобовые культуры, лен, конопля, морковь, виноград и др.; 4) Переднеазиатский — родина мягкой пшеницы, ячменя, овса; 5) Среднеземноморский — родина капусты, свеклы, маслин; б) Абиссинский — родина твердой пшеницы, сорго, бананов, кофе; 7) Центральноамериканский — родина кукурузы, какао, тыквы, табака, хлопчатника; 8) Южноамериканский — родина картофеля, ананаса, хинного дерева.
Дальнейшие исследования ученых привели к установлению еще четырех центров; Австралийского, Африканского, Европейско-Сибирского и Североамериканского (рис. 94).
Закон гомологических рядов наследственной изменчивости.
На примере злаков Н. И. Вавилов показал, что сходные признаки наблюдаются у разных видов данного семейства. Так, у пшеницы, ячменя, овса и кукурузы бывает белая, красная и черная окраска зерновок, существуют голые и пленчатые зерновки, встречаются колосья с длинными и короткими остями, безостые и с вздутиями вместо остей. В ходе последующих наблюдений было выяснено, что данный закон применим не только для растений, но распространяется на животных и микроорганизмы. Так, альбинизм встречается у всех классов позвоночных животных, короткопалость наблюдается у всех пород крупного рогатого скота, овец и собак.
Основные методы селекции растений.
Биологические особенности растений позволяют в селекционной работе с ними использовать инбридинг, полиплоидию, искусственный мутагенез, отдаленную гибридизацию и другие методы.
Отбор и гибридизация являются основными и традиционными методами селекции растений. Применяя массовый или индивидуальный отбор, селекционер не создает ничего нового, а выделяет растения с полезными качествами, уже имеющиеся в популяции. Этим методом выведены многие сорта, в том числе так называемые сорта народной селекции, например знаменитый по своим качествам сорт яблони Антоновка.
Для создания сортов растений с запрограммированными качествами ведется специальная целенаправленная работа — подбирается исходный материал, проводится гибридизация с последующим отбором.
Используя метод гибридизации с последующим отбором, селекционеры получили ценные высокоурожайные сорта пшеницы, ржи, подсолнечника, овощных, плодовых и других культур.
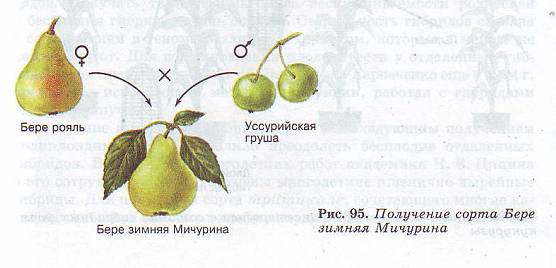
В разработку теории и практики селекции растений большой вклад внес ученый-селекционер Иван Владимирович Мичурин (1855— 1935), Он вывел около 300 новых сортов плодовых растений. В своих работах он широко применял скрещивание географически отдаленных форм. Так, скрещивая французский сорт груши Бере рояль с дикой уссурийской и выращивая сеянцы в условиях средней полосы России, он создал сорт Бере зимняя, сочетающий высокие вкусовые качества плодов с зимостойкостью (рис, 95). Методы, разработанные И. В. Мичуриным, успешно используются селекционерами и в настоящее время.
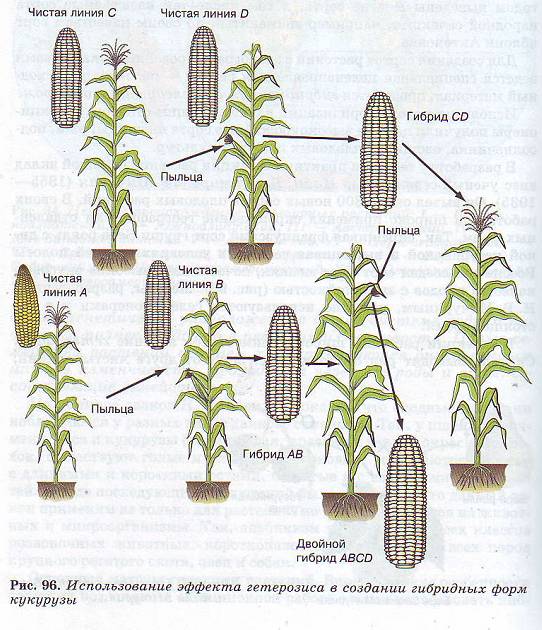
В селекции растений широко применяется явление гетерозиса.
Сначала выводят ряд отличающихся друг от друга чистых линий, а затем производят межлинейное скрещивание.
Выяснив, в каких случаях эффект гетерозиса проявляется наиболее сильно, используют лишь эти линии для получения гибридных семян. Эта методика применяется для получения высоких урожаев кукурузы, огурцов, томатов и других культур (рис. 96).

Полиплоидию (кратное увеличение числа хромосом) издавна использовали при создании сортов пшеницы, овса, картофеля, хлопчатника, плодовых, декоративных и других культур. Полиплоидные растения появлялись в популяциях случайно в результате естественных мутаций. В настоящее время применяют методы искусственного получения полиплоидов, воздействуя на растения разными мутагенами (в основном колхицином), разрушающими веретено деления клетки. Таким образом из диплоидных (2n) можно получить тетраплоидные (4n) формы.
Большинство их неперспективны, но отдельные формы служат ценным материалом для гибридизации и отбора. Полиплоидные растения могут отличаться более крупными размерами, высокой урожайностью и более активным синтезом органических веществ. Использование метода полиплоидии позволило селекционерам получить ценные сорта сахарной свеклы, ржи, гречихи, фасоли и других культур (рис. 97).
Отдаленная гибридизация позволяет в одном организме совместить признаки, характерные для растений разных видов и даже родов. Получать такие формы из-за нескрещиваемости родителей и бесплодия гибридов очень сложно. Стерильность гибридов связана с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. Для восстановления плодовитости у отдаленных гибридов известный генетик Георгий Дмитриевич Карпеченко еще в 1924 г. предложил использовать метод полиплоидии, работая с гибридами редьки и капусты.
Сочетание отдаленной гибридизации с последующим получением полиплоидных форм позволило преодолеть бесплодие отдаленных гибридов. В результате многолетних работ академика Н. В. Цицина и его сотрудников были получены многолетние пшенично-пырейные гибриды. Для получения сорта тритикале, сочетающего многие качества пшеницы (высокие хлебопекарные качества) и ржи (высокое содержание незаменимой аминокислоты лизина, а также способность расти на бедных песчаных почвах), применялась следующая схема:
Р: пшеница (2n = 42) х рожь (2n = 14)
G : n = 21
F1 2n = 28 (все непарные)
G: Мейоз нарушен, гибрид стерилен, нормальных гамет нет.
Обработка колхицином приводит к удвоению числа хромосом,
F1 :(колхицированное): 2n = 56
G : n = 28
F2, F3, , Fn : 2n= 56 (тритикале)
У таких гибридов в клетках содержится полный диплоидный набор хромосом обоих родителей, поэтому их хромосомы конъюгируют друг с другом и мейоз проходит нормально.
С помощью метода отдаленной гибридизации с последующим получением полиплоидных форм были выведены новые перспективные сорта картофеля, табака и других культур.
Методами отдаленной гибридизации и радиационного мутагенеза созданы перспективные сорта хлопчатника. Химический мутагенез лежал в основе получения многих новых сортов кукурузы, пшеницы, риса, овса, подсолнечника.
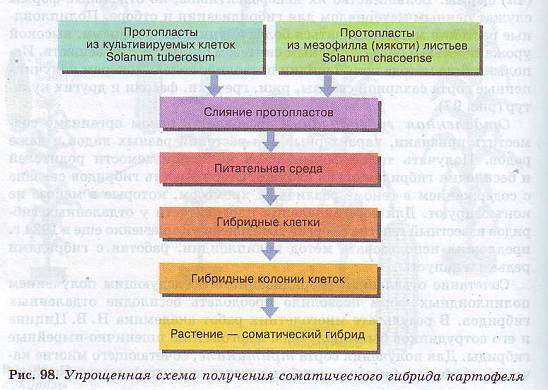
Методы клеточной инженерии.
Селекционеры все шире начинают применять для получения новых сортов растений методы клеточной инженерии. В качестве примера можно привести работу по соматической гибридизации двух видов картофеля: культурного — Solanum tuberosum и дикого Solanum chacoense (рис. 98). Для гибридизации использовались протопласты (греч. protos — первый и греч. plastos — вылепленный, образованный) — клетки, полностью лишенные клеточной стенки (оболочки) и имеющие только клеточную мембрану, которая ограничивает цитоплазму с различными органоидами.
Полученный соматический гибрид в сравнении с родительскими формами имел промежуточные характеристики по форме листа, величине клубней, но отличался большей мощностью куста и высотой стеблей, благодаря чему и был включен в дальнейшую практическую селекционную работу (рис. 99).

Метод вегетативного размножения культурой тканей широко применяется в селекции для быстрого размножения новых перспективных сортов растений.
В различных регионах нашей страны созданы научно-исследовательские институты и селекционные станции, которые проводят работы по выведению и районированию новых сортов растений. Эта работа играет важнейшую роль в повышении урожайности сельскохозяйственных культур и обеспечении населения продовольствием.
Центры происхождения культурных растений. Закон гомологических рядов наследственной изменчивости.
1. Какие методы применяются в селекции растений?
2. Какое значение для селекции имеет открытие закона гомологических рядов наследственной изменчивости?
3. Почему межлинейные гибриды сохраняют ценные признаки при вегетативном размножении и теряют их при семенном?
4. Почему селекционеры стремятся Получить растения-полиплоиды?
5. Какая методика позволяет преодолеть стерильность межвидовых (межродовых) гибридов?
Онлайн библиотека с учениками и книгами, плани-конспекти уроковс Биологии 10 класса, книги и учебники согласно календарного плана планирование Биологии 10 класса
Если у вас есть исправления или предложения к данному уроку, напишите нам.
Если вы хотите увидеть другие корректировки и пожелания к урокам, смотрите здесь - Образовательный форум.
Естественные автотетраплоиды возникают при слиянии гамет с нередуцированным (2n) числом хромосом, искусственные получают путем обработок колхицином. По сравнению с диплоидами автотетраплоиды характеризуются мощным ростом и большим размером частей растения. Последнее является следствием увеличения числа хромосом и объема клеток.
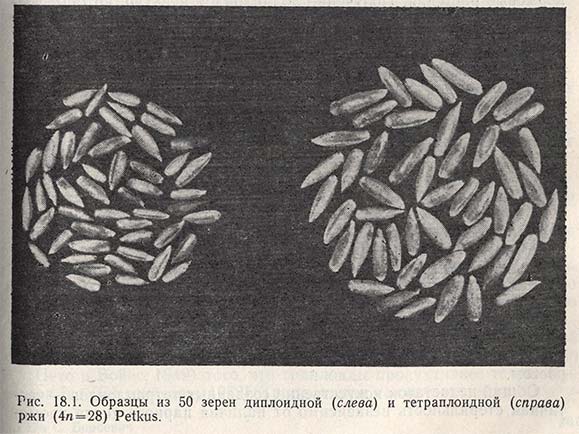
Тетраплоидная рожь (4n = 28). Первые тетраплоиды ржи получил Дорси, используя температурный шок; позднее (1941 г.) Мюнцинг использовал для этого колхицин. Растения тетраплоидной ржи имеют толстый стебель, широкие листья, длинные колосья, но особенного успеха удалось достигнуть по массе зерна, которая почти в два раза превышает таковую у диплоидов (рис. 18.1). С возрастанием крупности семян изменяется и соотношение углеводов и белка, причем тетраплоидная рожь содержит приблизительно на 2% больше белка и обладает более высокими хлебопекарными качествами, чем диплоидная. Серьезным недостатком тетраплоидной ржи является высокий процент стерильности колоса, на 20% превышающей стерильность диплоидной ржи в полевых условиях. Вследствие этого число зерен в колосе у нее меньше и получить значительную прибавку урожая по сравнению с диплоидной рожью не удается. Стерильность тетраплоидной ржи - это результат образования тривалентов и унивалентов, ибо еще не всегда удается восстановить необходимую сбалансированность между хромосомами и цитоплазмой.
Из-за довольно высокой стерильности тетраплоидная рожь не получила широкого распространения в производстве. Поэтому необходимо постоянно вести отбор лучших колосьев и создавать новые тетраплоидные сорта, скрещивание которых может привести к снижению стерильности. Медленное внедрение тетраплоидной ржи связано и с тем, что рядом с ней нельзя располагать посевы диплоидной ржи: это ведет к взаимному переопылению и возникновению полностью стерильных триплоидных (3n) растений. Тем не менее тетраплоидную рожь с успехом используют для выращивания на зеленый корм, так как она развивает мощные вегетативные органы.
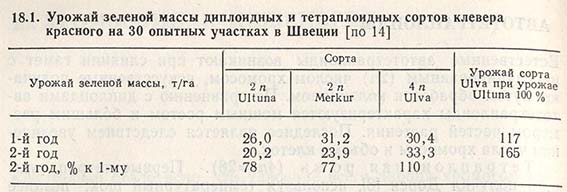
Тетраплоидный клевер красный (4n = 28) и другие кормовые растения. Получение полиплоидов культур, которые выращивают главным образом из-за зеленой массы, представляет большую практическую ценность. Так, в производстве уже находятся тетраплоидный клевер красный (табл. 18.1), вика и некоторые другие виды кормовых культур.
Ввиду значительных изменений и нарушений во взаимоотношениях между ядром и цитоплазмой цикл прохождения митоза у тетраплоидов обычно замедлен, вследствие чего они часто позднее созревают, чем диплоиды. Точно так же количество фаз митоза может быть меньше, и наконец, несмотря на более мощное развитие органов растений, тетраплоиды обычно не отличаются сильным ростом.
Общий недостаток искусственно созданных тетраплоидов - частичная стерильность независимо от наличия парного числа хромосом. Автотетраплоиды имеют четыре гомологичные хромосомы (диплоиды - две), которые в пахитене образуют квадриваленты. Если в анафазе I одна пара хромосом отходит к одному полюсу, а другая - к другому, то половые клетки содержат одинаковое число хромосом и способны к оплодотворению. Однако иногда хромосомы образуют не квадриваленты, а триваленты и униваленты. В результате появляются гаметы с неодинаковым числом хромосом, а гаметы, содержащие неполный гаплоидный набор хромосом, не способны к оплодотворению. По всем этим причинам и возникает частичная стерильность, которая снижает практическую ценность известного числа тетраплоидов.
Читайте также: