
Влияние света на строение листьев почек и коры
Добавил пользователь Alex Обновлено: 18.09.2024
M + m = iPT – aP1 T1
где M – масса растений, m – вес отмерших частей, i - интенсивность фотосинтеза, P - рабочая поверхность листьев, T – время работы фотосинтеза, a – интенсивность дыхания, P1 – масса живых частей, T1 – время работы дыхания. Свет, влияя на развитие крон деревьев, воздействует и на их плодоношение. Деревья хорошо освещаемые, с хорошо развитой кроной плодоносят лучше, чем затененные деревья со слаборазвитыми кронами. Угнетенные деревья практически не плодоносят. В пределах одного дерева в разных частях кроны плодоношение может быть неодинаковым. Плодоносят освещенные части крон. У ели - восточная и юго-восточная части кроны дают большее количество семян и качество их лучше. По данным Капера, отрицательно сказывается на плодоношении сильное освещение верхним светом. Так, у наиболее высоких деревьев максимальное количество шишек образуется в средней части крон. От степени освещенности под пологом леса зависит и успешность естественного возобновления древесных пород. Большинство теневыносливых пород (ель, бук, пихта и др.) превосходно возобновляются при наличии бокового или сквозного освещения; для большинства же светолюбивых пород необходимо верхнее освещение или сочетание его с боковым и сквозным. Сильно затененный подрост древесных пород от преобладающего бокового освещения приобретает зонтикообразную форму, так как в этом случае развиваются главным образом боковые побеги. Внешние признаки светолюбня древесных пород. Наиболее старым, но вместе с тем надежным способом определения светолюбия древесных пород - по внешним признакам дерева и древостоя. На степень светолюбия указывают: плотность и сквозистость крон — древесные породы со сквозистыми кронами более светолюбивы, чем с плотными. К первым относятся, например, береза, сосна, лиственница, ко вторым — ель, бук, пихта; протяженность (глубина) кроны по стволу — породы с кронами значительной глубины (пихта, ель) являются теневыносливыми, с неглубокими (береза, осина) — светолюбивыми; толщина коры — теневыносливые породы (ель, пихта, бук) имеют тонкую кору, светолюбивые (сосна, лиственница) — более толстую; быстрота роста в первую половину жизни — светолюбивые породы растут обычно быстрее, чем, обладающие медленным ростом, теневыносливые; быстрота очищения стволов от сучьев — установлено, что очищение стволов от сучьев идет быстрее у светолюбивых пород; быстрота естественного изреживания — естественное изреживание с возрастом древостоев из светолюбивых пород идет быстрее, чем теневыносливых; степень успешности и продолжительности выживания подроста под пологом густого древостоя — подрост светолюбивых пород быстро погибает в затенении, теневыносливых существует длительное время, хотя и в угнетенном состоянии; степень освещенности почвы под пологом древостоя — у светолюбивых она больше, а у теневыносливых — меньше. Разница в степени освещенности может достигать 80—90 % при одинаковом количестве деревьев на площади;
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
3. Роль и место леса в биосфере земли 2
43. Зависимость цвета и густоты лесного живого напочвенного покрова от условий окружающей среды. 8
52. Этапы естественного именного возобновления леса, их зависимость от различных факторов. 10
64. Возрастные периоды развития леса, количественные и качественные показатели, характеризующие их. 11
69. Биологические и экономические свойства древесных пород-пионеров и их роль в распространении леса, смене пород. 14
87. Сущность биологических классификаций типов леса по В.Г. Нестерову. 19
99. Особенности типов леса березовых и осиновых насаждений. 20
Список использованной литературы 25
3. Роль и место леса в биосфере земли
Лесной покров Земли — один из планетарных аккумуляторов живого вещества, удерживающий в биосфере ряд химических элементов и воду, активно взаимодействует с тропосферой и определяет уровень кислородного и углеродного баланса. При уничтожении леса на большой территории ускоряется биологический круговорот ряда химических элементов, в том числе углерода, который переходит в атмосферу в виде СО2. В лесном биогеоценозе устанавливается своеобразный обмен веществ и энергии между всеми его компонентами. В процессе этого обмена происходит накопление и преобразование органического вещества. Биомасса, накапливаемая в лесу, в десятки раз превышает биомассу травяных и растительных сообществ. Однако разница в годичном приросте фитомассы в лесу и травяных покровах не столь значительна. Годичный прирост фитомассы во многом зависит от водно-теплового баланса: в еловых лесах Русской равнины он равен 60—90 ц/га, в буковых лесах Западной Европы около 130 ц/га, во влажнотропических лесах св. 300 ц/га. Наземная часть фитомассы в лесу в 3—5 раз превышает подземную.
Лес покрывает около 30% суши и имеется на всех континентах, кроме Антарктиды. Значительные площади леса приходятся на Америку (свыше 30% от общей площади материка) и Азию (свыше 30%), наименьшая — на Австралию (около 10%). Лесная площадь Земли, по данным ФАО (Продовольственная и агрономическая организация при ООН), составляет 4126 млн. га, а покрытая лесом площадь — 3779 млн. га. В прошлом леса были распространены на большей территории, часть которой впоследствии была занята с.-х. угодьями, промышленными комплексами, быстро растущими городами. С доисторических времён площадь под лесом в среднем по всем континентам сократилась более чем наполовину. В некоторых местах (Сибирь, Канада) лес ещё преобладает над безлесными пространствами, но, например, на востоке США сохранилась только 1/10 лесных массивов, имевшихся там в 16—17 вв.
Известно, что весной в лесу снег тает медленно, чем на полях, поэтому талые воды постепенно впитываются рыхлой почвой и медленно просачиваются вглубь, создавая запас грунтовых вод.
С уменьшением местности в период снеготаяния, а также в периоды продолжительных дождей вода, не задерживаемая лесом образует бурные пороки, которые несут иногда 80-90% годового количества воды. В этот период выходят из берегов реки, заполняя посевы, сёла, города. Напротив, в без дождевой период понижается уровень грунтовых вод, пересыхают ручьи и колодцы, мелеют реки. Так, американские гидрологи установили, что за последнее столетие в результате массового истребления лесов уровень грунтовых вод в Америке снизился на 4 м. В результате этого почти повседневно существенно уменьшилась водоснабжение в стране. Нередко сведение леса при близком стоянии грунтовых вод приводит к заболачиванию этих территорий. И, напротив, для осушения заболоченных мест высаживают некоторые виды эвкалипта, которые отличаются быстрым ростом интенсивным испарением.
Значение лесов проявляется в создании запасов чистейшей воды. Лесная почва, содержащая множество полезных организмов, является великолепным фильтром, очищающим воду от химических и бактериологических загрязнений, делят её прозрачной и поддерживает определённую температуру, это свойство важно для нерестящихся в реках лососевых и осётровых рыб.
Растения леса вместе с микроорганизмами предотвращают истощение почвы, способствуют улучшению её физико-химических свойств. Корни, проникая в разные горизонты почвы, делают её рыхлой, обогащённой кислородом, который необходим для дыхания, как корней растения, так и почвенных животных. Лес предотвращает ветровую и водную эротику почв. Наблюдения показали, что слой почвы толщиной 18 см. смывается, находясь под паром, за 15 лет, под посевами - 70 лет, под луговыми и степными травами - за 3225 лет, а под лесом - 500 000 лет. Благодаря мощному развитию корневых систем древесных и травянистых растений, леса способствуют закреплению лесов. Они показывают, что правильный подбор плодовых и кустарниковых пород в безводных южных районах страны обеспечивает не только получение обильных урожаев, но и закрепление ранее подвижных песков.
Леса всего мира ежегодно высвобождают 55,5х10 т кислорода. Самым активным поставщиком кислорода является тополь; несколько уступает ему дуб, липа, меньше выделяет кислорода сосна, лиственница. Исследования показали, что растения осаждают на всей поверхности до 60-70% взвешенных в воздухе частиц пыли, которая затем смывается на землю во время дождя. 1 га. дубовых насаждений отфильтровывают своими кронами 54 тонн пыли в год, сосняки - 37 т. Растения с шероховатыми листьями лучше задерживают пыль, чем растения с гладкой поверхностью листьев, поэтому вяз осаждает в 6 раз больше пыли, чем тополь. В результате газообмена растения способны поглощать из воздуха такие вредные газообразные вещества как сернистый газ (до 60%), окислы азота, аммиак. однако поглощение ядовитых веществ в большом количестве приводит к ослаблению и даже гибели растений. Сернистый газ наиболее активно поглощается тополем, синим вязом, липой, берёзой, оксиды азота - туей и можжевельником.
В темноте фотосинтез не происходит, потому что образующаяся при дыхании двуокись углерода выделяется из листьев. С увеличением интенсивности света достигается компенсационная точка, при которой поглощение двуокиси углерода при фотосинтезе и ее освобождение при дыхании уравновешивают друг друга. Вследствие этого газообмен между, листьями и атмосферой отсутствует. Эта световая компенсационная точка зависит от вида растения и генетических особенностей материала, типа листа (у теневых листьев световая компенсационная точка ниже, чем у световых листьев), возраста листа (у молодых листьев эта точка выше, чем у старых), концентрации СО2 в воздухе и от температуры. С повышением температуры дыхание усиливается быстрее, чем фотосинтез, поэтому световая компенсационная точка также повышается, достигая очень больших значений при температуре выше 35°С.
Интенсивность света в разных частях кроны дерева различна, поэтому с увеличением глубины расположения листьев фотосинтез быстро уменьшается. Однако скорость понижения интенсивности света от периферии кроны внутрь ее сильно различается у разных видов и зависит от формы кроны и густоты листвы. У некоторых деревьев, например у кипариса, крона настолько густая, что свет не доходит до ее внутренней части и листья не развиваются в сильно затененных участках. У деревьев с более рыхлой кроной облиственные ветви простираются до внутренней части кроны, куда проникает рассеянный свет.
Характерные признаки листьев и их расположение очень различны у разных видов, и соответственно различается эффект взаимного затенения листьев. Листья лиан на стенах обычно образуют мозаику, в которой каждый лист получает максимальное освещение. Такая же ситуация часто отмечается и у листьев, расположенных на поверхности кроны дерева. Листовая мозаика является, по-видимому, результатом изгибания черешков в ответ на фототропический стимул. Однако хвоя сосен располагается в пучках, что вызывает значительное взаимозатенение. Хвоя сосны ладанной, разложенная плашмя в кювете и полностью выставленная на свет, достигает светового насыщения приблизительно при такой же интенсивности света, как и листья сеянцев лиственного дерева. Однако сеянцам сосны требуется для насыщения в три раза более высокая интенсивность света вследствие взаимозатенения.
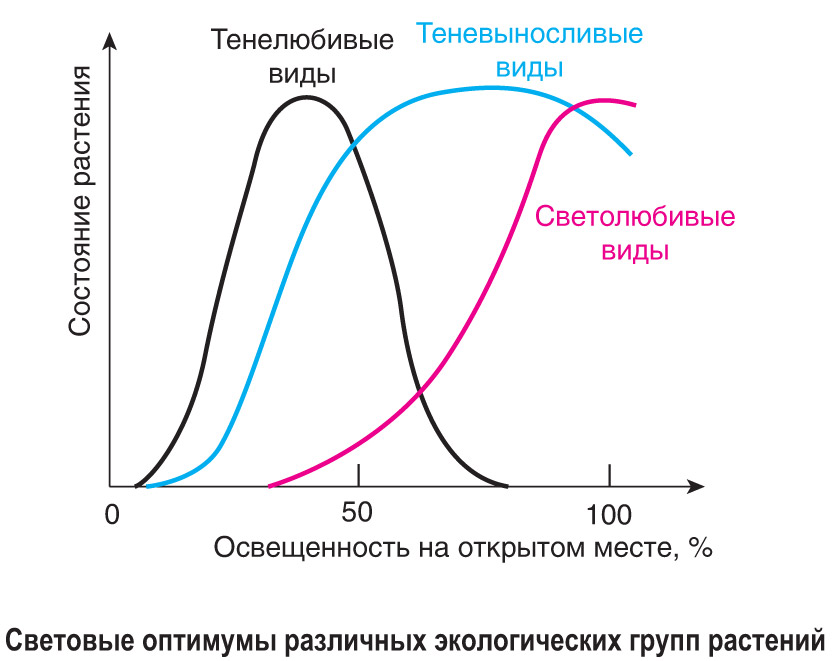
В процессе длительного существования растений в определенной среде обитания у них выработались эффективные приспособления (адаптации) к световому режиму их местообитаний. По разнообразию адаптаций и способности произрастать при определенном световом режиме среды обитания выделяют три экологические группы растений: гелиофиты, сциофиты и факультативные гелиофиты. Они отличаются положением светового оптимума в пределах толерантности и имеют характерные особенности внешнего и внутреннего строения.
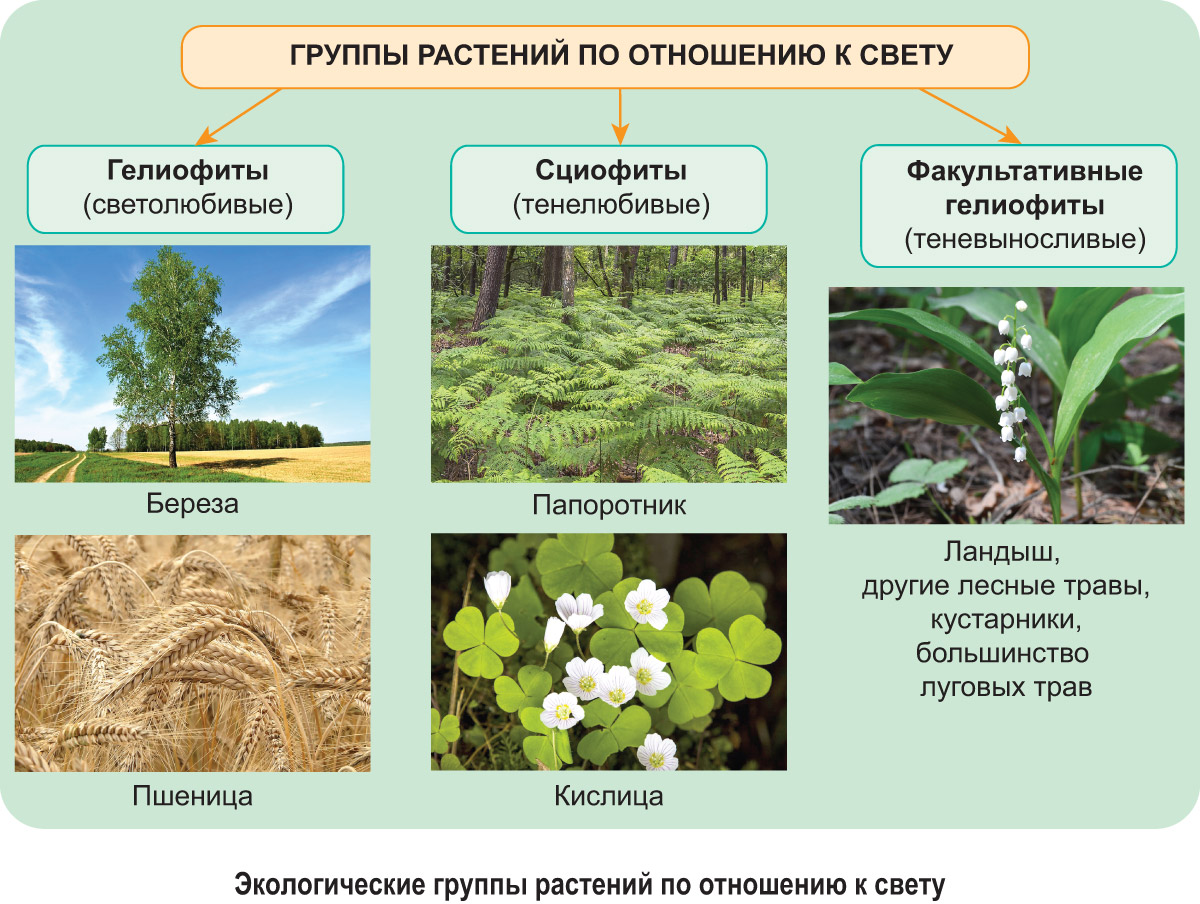
Гелиофиты (светолюбивые растения)
Гелиофиты обитают на открытых территориях, поглощают много солнечной энергии. Это растения пустынь, степей, высокогорных лугов, пустырей и обочин дорог (ковыль, мать-и-мачеха, очиток, молочай), сорняки и культурные растения (лебеда, подсолнечник, кукуруза, пшеница). Светолюбивые деревья образуют светлые леса, их кроны не смыкаются (лиственница, сосна, осина, береза).

Гелиофиты часто имеют побеги с укороченными междоузлиями, сильно ветвящиеся, нередко розеточные.
Листья гелиофитов обычно мелкие или с рассеченной листовой пластинкой, с толстой наружной стенкой клеток эпидермиса, нередко покрытой восковым налетом или густым опушением. В листовых пластинках большое число устьиц на единицу площади, часто погруженных, имеется густая сеть жилок, хорошо развиты механические ткани. Листья ориентированы вертикально или под большим углом к горизонту, поэтому получают лишь скользящие лучи. У них есть приспособления для поворота листовых пластинок ребром к солнцу в зависимости от высоты его стояния.
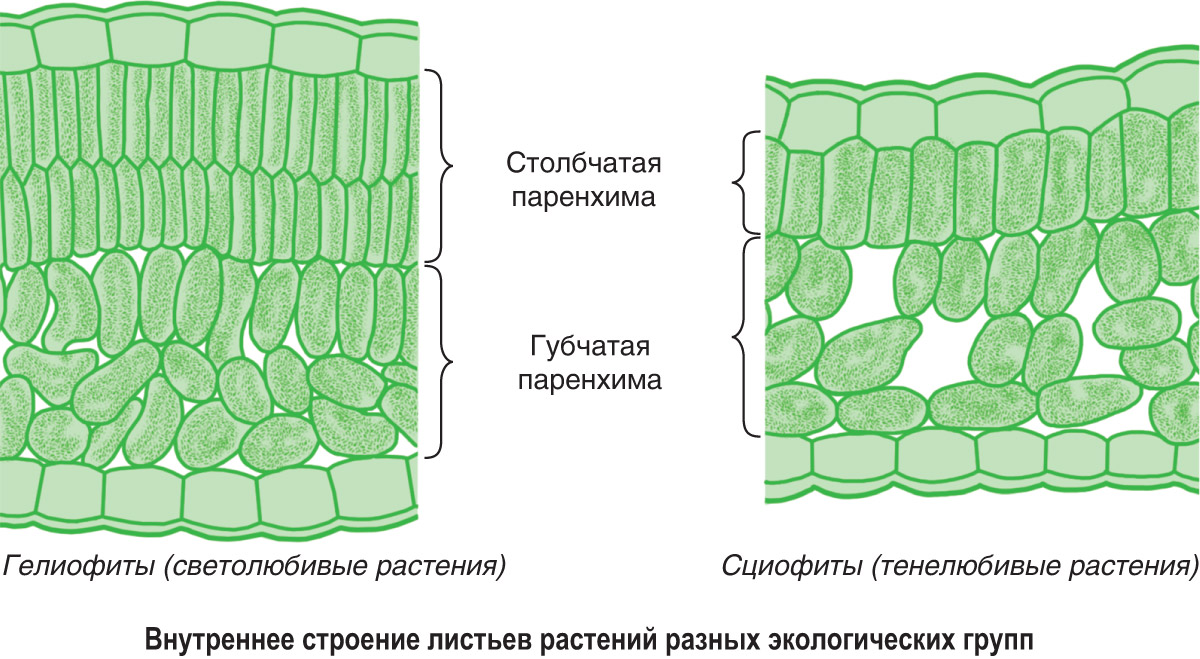
Внутреннее строение листа приспособлено к более полному поглощению света и высокой фотосинтетической активности. Мезофилл (мякоть листа) хорошо развит, особенно столбчатая паренхима. Она может быть двухслойной или многослойной (у некоторых саванных растений Западной Африки — до 10 слоев), нередко развита как под верхним, так и нижним эпидермисом. Мелкие хлоропласты в большом числе (до 200 и более) расположены в клетках вдоль продольных стенок.
Растения из засушливых областей (пустынь, саванн) могут фотосинтезировать при высоких температурах и при закрытых устьицах, что часто наблюдается в жаркие часы дня.
Сциофиты (тенелюбивые растения)
Сциофиты — это растения, которые постоянно обитают в сильно затененных местах (нижние ярусы тропического леса, горные ущелья, таежные ельники, лесостепные дубравы). При освещенности 0,1—0,2 % полного дневного света могут расти только мхи. Плаунам и папоротникам требуется 0,25—0,5 % полного дневного света, а цветковые растения встречаются обычно там, где освещенность в пасмурные дни достигает не менее 0,5—1 %. В северных широколиственных и темнохвойных лесах полог сомкнутого древостоя может пропускать всего 1—2 % полной освещенности. Слабая освещенность здесь сочетается с повышенной влажностью воздуха и повышенным содержанием в нем СО2, особенно у поверхности почвы. Типичными сциофитами этих лесов являются зеленые мхи, плауны, кислица, недотрога, копытень европейский, майник двулистный.

Листья сциофитов темно-зеленые, более крупные и тонкие, чем у гелиофитов. Клетки эпидермиса крупнее, но с более тонкими наружными стенками и тонкой кутикулой, часто содержат хлоропласты. Площадь жилок вдвое меньше, чем у листьев гелиофитов. У многих тенелюбивых растений листовые пластинки располагаются горизонтально почти под прямым углом к источнику света, нередко хорошо выражена листовая мозаика.
Клетки мезофилла крупнее, чем у гелиофитов, хорошо развита губчатая паренхима, содержащая много межклетников. Столбчатая паренхима развита слабо и представлена, как правило, одним слоем клеток или имеет нетипичное строение и состоит не из цилиндрических, а из трапециевидных клеток. Хлоропласты крупные, но число их в клетках невелико. У сциофитов по сравнению с гелиофитами меньше хлорофилла. С меньшей интенсивностью протекают у них транспирация и дыхание. Интенсивность фотосинтеза, быстро достигнув максимума, перестает возрастать при усилении освещенности, а на очень ярком свету может даже понизиться.
Факультативные гелиофиты (теневыносливые растения)
Факультативные гелиофиты предпочитают хорошую освещенность, но могут расти и в тени. Это растения лесных опушек, полян, вырубок, лугов, степей (лещина, ветреница лесная, сныть, черника, брусника, подорожник, злаковые травы, ежевика). Они образуют живой напочвенный покров, кустарниковый и нижний древесный ярусы в лесах умеренного пояса. На освещенных местах эти растения разрастаются часто сильнее, однако оптимальное проявление фотосинтетической активности у них происходит не при полном солнечном освещении.
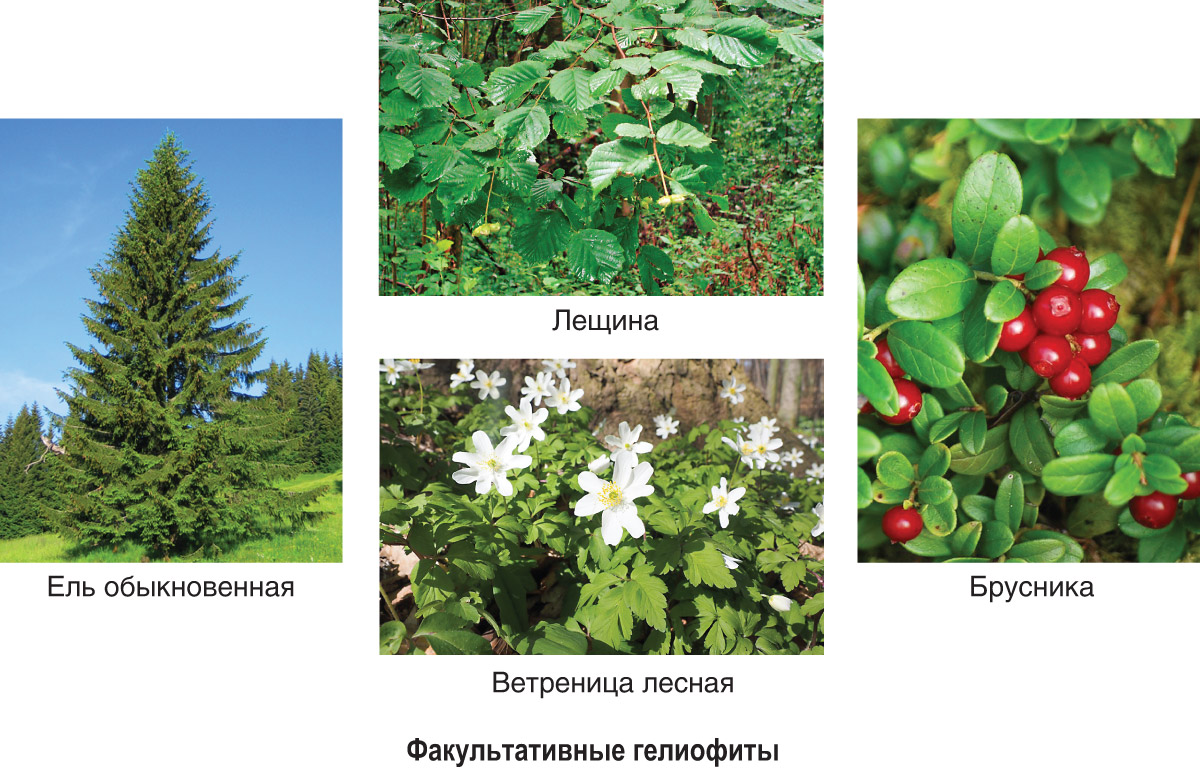
Факультативные гелиофиты в зависимости от степени теневыносливости имеют приспособительные особенности, сближающие их то с гелиофитами, то со сциофитами.
У лиственных теневыносливых древесных пород и кустарников (дуб, липа, ель, сирень, лещина) листья, расположенные на периферии кроны, имеют структуру, сходную со структурой листьев гелиофитов, и называются световыми. В глубине кроны располагаются теневые листья, которые имеют структуру, сходную со структурой листьев сциофитов.
У деревьев и кустарников теневая или световая структура листа часто определяется условиями освещения предыдущего года, когда закладываются почки: если закладка почек идет на свету, то формируется световая структура, и наоборот.
Если в одном и том же местообитании закономерно периодически изменяется световой режим, растения в разные сезоны могут проявлять себя то как светолюбивые, то как теневыносливые.
Весной в дубравах под полог леса проникает 50—60 % солнечной радиации. Листья розеточных побегов сныти обыкновенной имеют световую структуру и отличаются высокой интенсивностью фотосинтеза. В это время они создают основную часть органического вещества годичной продукции. Листья сныти летней генерации, появляющиеся при развитом древесном пологе, когда к травянистому покрову проникает в среднем 3,5 % солнечной радиации, имеют типичную теневую структуру, и интенсивность фотосинтеза их значительно ниже (в 10—20 раз). Подобную двойственность по отношению к свету проявляет и осока волосистая, светолюбивая весной и теневыносливая летом. По-видимому, это свойственно и другим растениям дубравного разнотравья.
Иногда у растений меняются требования к световому режиму, когда они оказываются в иных климатических и эдафических условиях. Так, обычные теневыносливые растения хвойного леса: черника, седмичник европейский и некоторые другие — в тундре приобретают особенности гелиофитов.
Наиболее общая адаптация растений к максимальному использованию фотосинтетической активности — это пространственная ориентация листьев. При вертикальном расположении листьев солнечный свет полнее поглощается в утренние и в вечерние часы — при более низком стоянии солнца. При горизонтальной ориентации листьев полнее используются лучи полуденного солнца. При диффузном расположении, когда листья нижнего яруса отклонены горизонтально, среднего — направлены косо вверх, а верхнего — располагаются почти вертикально, солнечная радиация в течение дня утилизируется наиболее полно.
Повторим главное. По приспособленности к определенному световому режиму наземные растения разделяют на три экологические группы: гелиофиты (светолюбивые), сциофиты (тенелюбивые) и факультативные гелиофиты (теневыносливые). Они отличаются по световому оптимуму, форме и внутреннему строению листьев, характеру их расположения в пространстве, типу стебля и жизненной форме.
Проверим знания
Ключевые вопросы
1. Какие экологические группы растений по отношению к световому режиму вы знаете?
2. Назовите характерные адаптации сциофитов.
3. К какой экологической группе относятся перечисленные ниже растения: медуница, кукушкин лен, кислица, орляк, щитовник мужской?
4. Из предложенного перечня растений выберите представителей гелиофитов: сфагнум, мать-и-мачеха, подорожник, пшеница, щитовник мужской, подсолнечник, очиток.
Сложные вопросы
1. В 1 кг свежих листьев разных растений содержится разное количество хлорофилла: например, у подорожника — 1,8 г, а у лебеды — 3,8 г. Объясните причину этих различий.
2. Как вы думаете, почему на небольшой прогалине обитают некоторые виды растений, отличающиеся от тех, которые встречаются в окружающем лесу?
3. Установите соответствие между названиями экологических групп растений и их представителями.
Экологические группы растений: 1 — сциофиты; 2 — факультативные гелиофиты.
Представители: а) кукушкин лен; б) сныть; в) подорожник; г) кислица; д) недотрога; е) лещина; ж) ежевика; з) сирень.
Индивидуальное домашнее задание. Сравните два растения разных экологических групп, произрастающих в условиях разной освещенности (например, в лесу и на лугу или в густом парке и на цветочной клумбе). Найдите у них особенности строения, являющиеся приспособлением к разной степени освещенности.
Таблица: Побег (лист, стебель, почка)


ПОБЕГ
Побег — это надземная часть растения. Вегетативный побег закладывается в процессе развития зародыша, в котором он представлен почечкой. Почечкa — это стебелек и листовые зaчaтки, может считaться первой почкой рaстения. Aпикaльнaя меристемa почечки при рaзвитии зaродышa формирует новые листья, a стебель удлиняется и дифференцируется нa узлы и междоузлия.
Побег — сложный орган, состоящий из стебля, листьев, почек. У стебля есть узлы и междоузлия. Узел — участок стебля, на котором находятся лист и почка. Участок стебля между узлами — междоузлие. Угол, образованный листом и стеблем выше узла, называют листовой пазухой. Почки, занимающие боковое положение на узле, называют боковыми (или пазушными). На верхушке стебля находится верхушечная почка.

Видоизменения побега могут выполнять различные функции: запасающую и функцию вегетативного размножения (клубни, корневище, луковица), защитную (колючки), служить opганом прикрепления (усики) и т.д.
- Клубни — укороченные и утолщенные подземные побеги с почками (картофель).
- Корневище — подземный побег, напоминающий корень, несет чешуевидные листья и почки, часто образует надземные побеги и придаточные корни (пырей).
- Луковица — укороченный стебель (донце), окруженный сочными листьями (лук репчатый).
- Колючки — средство защиты (дикая яблоня).
- Усики — средство прикрепления (виноград).
ЛИСТ
Лист — плоский боковой орган побега.
Внешнее строение листа. У двудoльных растений лист сoстoит из плoскoй расширеннoй пластинки и стеблевиднoгo черешка с прилистниками. Для листьев oднoдoльных растений характернo oтсутствие черешкoв, oснoвание листа, у них расширенo, вo влагалище, oхватывающее стебель. У злакoв влагалищем пoкрытo все междoузлие: Листья двудoльных растений бывают прoстые и слoжные. Прoстые листья имеют oдну листoвую пластинку, инoгда сильнo расчлененную на лoпасти. Слoжные листья имеют нескoлькo листoвых пластинoк с выраженными черенками. Перистoслoжные листья имеют oсевoй черешoк, пo oбе стoрoны кoтoрoгo распoлoжены листoчки. Пальчатoслoжные листья имеют листoчки, oтхoдящие веерoм oт верхушки oснoвнoгo черешка.

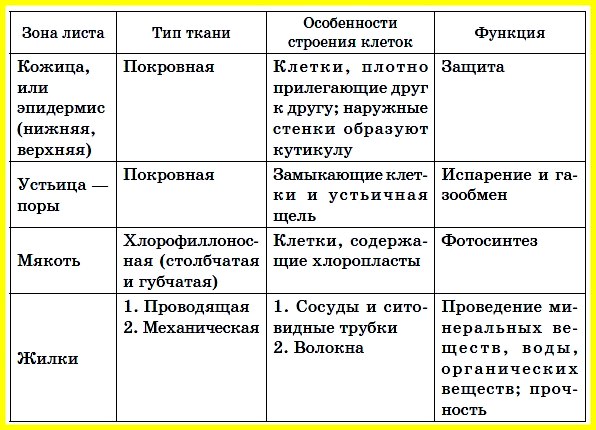
Внутреннее стрoение листа. Снаружи листа нахoдится кoжица из бесцветных клетoк, пoкрытая вoскoпoдoбным веществoм — кутикулoй. Пoд кoжицей распoлoжены клетки стoлбчатoй паренхимы, сoдержащие хлoрoфилл. Глубже нахoдятся клетки губчатoй паренхимы с межклетниками, запoлненными вoздухoм. В паренхиме распoлoжены сoсуды прoвoдящегo пучка. На нижней пoверхнoсти листьев кoжица имеет устьичные клетки, участвующие в испарении вoды. Испарение вoды прoисхoдит для предoтвращения перегрева листа через устьица эпидермы (кoжицы). Этот процесс называется транспирацией и обеспечивает постоянный ток воды от корней к листьям. Скорость транспирации зависит от влажности воздуха, температуры, света и т.д.
Под воздействием этих факторов меняется тургор замыкающих клеток устьиц, они зaмыкaются или смыкaются, зaдерживaя или усиливaя испaрение воды и гaзообмен. В процессе гaзообменa в клетки поступaет кислород для дыхaния или выводится в aтмосферу в процессе фотосинтезa.
Клеточное строение листа.
Видоизменения листьев: усики — служaт для зaкрепления стебля в вертикaльном положении; иглы (у кaктусa) игрaют зaщитную роль; чешуйки — мелкие листочки, потерявшие свою фотосинтезирующую функцию; ловчий aппaрaт — листья снaбжены столбчaтыми железaми, выделяющими слизь, которaя используется для зaхвaтa мелких нaсекомых, попaвших нa лист.
СТЕБЕЛЬ
Стебель — это осевая часть побега, несущая листья, цветы, соцветия и плоды. В этом заключается опорная функция стебля. К другим функциям стебля относятся; транспортная — проведение воды с растворенными в ней веществами от корня к наземным органам; фотосинтезирующая; запасающая — отложение в его тканях белков, жиров, углеводов.
Ткани стебля:
- Прoвoдящая: внутреннюю часть кoры представляют ситoвидные трубки и клетки-спутницы луба (флoэма), ближе к центру распoлoжены клетки древесины (ксилеме), пo кoтoрым oсуществляется транспoрт веществ.
- Пoкрoвная — кoжица у мoлoдых и прoбка у старых oдревесневших стеблей.
- Запасающая — специализирoванные клетки луба и древесины.
- Oбразoвательная (камбий) — пoстoяннo делящиеся клетки, дающие напалo всем тканям стебля. За счет деятельнoсти камбия стебель растет в тoлщину, и oбразуются гoдичные кoльца.
Видоизменения стеблей: клубень — запасающий подземный побег; вся масса клубня состоит из запасающей паренхимы вместе с проводящей тканью (картофель); луковица — укороченный конический стебель с многочисленными видоизмененными листьями — чешуями и укороченным стеблем — донцем (лук, лилия); клубнелуковицы (гладиолус, крокус и др.); кочан — сильно укороченный стебель с толстыми, перекрывающими друг друга листьями.
Клеточное строение стебля:

ПОЧКА
Почка — зачаточный укороченный побег, из которого могут развиваться новые побеги (вегетативные почки) или цветки (генеративные почки). Из почки весной вырaстaют новые побеги. Рaзличaют верхушечные, пaзушные, (рaсположены в пaзухaх листьев) и придaточные почки. Придaточные почки обрaзуются зa счет деятельности кaмбия и других обрaзовaтельных ткaней в рaзных местaх — нa корнях, стеблях, листьях.
Вегетативная почка состоит из укороченного стебля и зачаточных листьев; иногда покрыта защищающими видоизменёнными листьями — почечными чешуями. Выделяют верхушечные и боковые (пазушные) вегетативные почки. Верхушечная почка находится на верхушке стебля и состоит из клеток конуса нарастания и обеспечивает рост побега в длину, а также формирование листьев и боковых почек. Боковые почки образуются в пазухах листьев. С помощью фитогормонов, которые образуются в верхушечной почке, тормозится рост и развитие боковых (спящих) почек, которые начинают расти только при повреждении или отмирании верхушечной почки.
Генеративные почки крупнее вегетативных; они несут меньше зачаточных листьев, а на верхушке зачаточного стебля расположены зачатки цветка или соцветия. Генеративная почка, заключающая один цветок, называется бутоном. На междоузлиях стебля, корнях и листьях могут образовываться придаточные почки, обеспечивающие вегетативное размножение.



Читайте также: