
В состав ксилемы голосеменных и покрытосеменных растений входят сосуды ситовидные трубки трахеиды
Добавил пользователь Евгений Кузнецов Обновлено: 19.09.2024
Проводящие ткани осуществляют функцию проведения воды и растворов различных питательных веществ по всему телу растения. Они состоят из ксилемы (древесины), проводящей воду и растворённые в ней вещества из корней, и флоэмы (луба), проводящей из листьев органические вещества.
Ксилема (древесина)
В состав ксилемы (рис. 12) входят проводящие элементы — сосуды и трахеиды, а также живые паренхимные клетки и механические волокна.
Трахеиды
Трахеиды представляют собой замкнутые удлинённые клетки с вытянутыми концами и с утолщёнными одревесневшими стенками, на которых имеются поры. Передвижение растворов происходит через поры. Кроме проводящей функции трахеиды несут механическую нагрузку. У папоротников и голосеменных трахеиды служат единственным проводящим элементом.
Сосуды (трахеи)
Сосуды, или трахеи, представляют собой трубки, состоящие из многих клеток, поперечные перегородки между которыми разрушаются (рис. 13). Боковые стенки их утолщаются и одревесневают, но в них остаются поры, через которые происходит передвижение веществ в горизонтальном направлении. По характеру утолщений стенок различают кольчатые, спиральные, лестничные, точечные и сетчатые сосуды. Кольчатые и спиральные сосуды эластичны, они могут вытягиваться, поэтому не препятствуют росту органов растений и появляются в них раньше других. Протопласты трахеид и сосудов отмирают.
 |
| Рис. 13. Разные типы утолщения стенок сосудов и поровости: 1 — кольчатый; 2 — 4 спиральные; 5 — сетчатый; 6 — лестничный; 7 — супротивная поровость; 8 — очередная поровость |
Флоэма (луб)
Флоэма состоит из ситовидных трубок, клеток-спутниц, лубяной паренхимы и лубяных волокон.
Флоэма и ксилема являются сосудистыми тканями трахеофитных растений, то есть в них расположены сосуды растений, которые осуществляют транспортировку сока в растительных организмах: папоротникообразных, голосеменных и покрытосеменных, сообщая корневую систему с лиственными структурами посредством стебля. Обе являются переносчиками сока.

Флоэма транспортирует переработанный сок (или органический сок, продукт фотосинтеза, где питательные вещества превращаются в глюкозу) и находится в самом внешнем слое стебля. Она транспортирует переработанный сок из хлорофильных частей (мест, где происходит фотосинтез) к живыми частями растения — областям, где глюкоза будет превращаться в энергию. Флоэма образована живыми клетками, вытянутыми и без ядра.

Транспортировка необработанного сока, образованного водой и минеральными солями, осуществляемая ксилемой, происходит из способности поглощения корней и распределения в основной пункт назначения к листьям. Ее основной состав включает сосуды, трахеи, трахеиды, волокна и паренхимные клетки.
Транспортировка переработанного сока, образованного органическими веществами, полученными при фотосинтезе, осуществляется через флоэму, начиная от листьев в направлении остальных органов, главным образом, энергетического резерва (корней и стеблей). Она состоит из ситовидных элементов, сопровождающих клеток, волокон и паренхимных клеток.
Растения как автотрофные организмы
У растений автотрофное питание, другими словами, они могут производить органические молекулы (главным образом глюкозу) из простых неорганических веществ с помощью внешнего источника энергии, энергии света.
Итак, питательные вещества, которые требуются растению, очень просты: вода, минеральные соли и углекислый газ (мы не называем солнечную энергию, потому что это не вещество, а тип энергии). Растения в процессе фотосинтеза, происходящего в хлоропластах клеток, продуцируют глюкозу, которая является органической молекулой, служащей источником химической энергии (а также для создания других малых и больших структурных молекул, например, целлюлозы).
В связи с этим растение испытывает следующие трудности:
- Фотосинтез происходит главным образом в клетках листьев на определенной высоте над землей (иногда несколько метров), но воду и минеральные соли следует добывать из почвы, корнями. Поэтому растение должно транспортировать эти простые вещества вверх, преодолевая силу тяжести.
- Не все клетки растения выполняют фотосинтез (например, клетки корней нет). Поэтому, как только глюкоза вырабатывается в клетках листьев, часть ее должна транспортироваться в другие места для снабжения всех клеток.
Растение должно поглощать воду и минеральные соли через корни. Для этого клетки самого поверхностного слоя корня имеют очень тонкие расширения в виде нитей, которые выступают наружу, называемые абсорбирующими или корневыми волосками. Таким образом, контактная поверхность корня с внешней средой значительно увеличивается.
Клетки, которые составляют абсорбирующие волоски, имеют важную способность: они способны переносить минеральные соли из внешней среды во внутреннюю часть клетки, даже если концентрация солей внутри клетки больше, чем снаружи (затратив некоторую энергию, конечно). Таким образом, концентрация солей становится больше внутри клеток по отношению к внешней стороне.
Существуют две альтернативы для проникновения воды в корень: вода может проходить через клетки, переходя из одной в другую через небольшие отверстия, называемые плазмодесмами (внутриклеточный или упрощенный транспорт) или проходить через пространства клеточных стенок, которые отделяют одну клетку от другой (внеклеточный или апопластический транспорт).
Как только вода была поглощена, достигнута первая цель: вода и минеральные соли находятся внутри корневых клеток… но все еще далеки от их цели, которая является листьями.
Транспортировка необработанного сока через ксилему

Для доставки воды, минеральных солей и других веществ, производимых корневыми клетками, к листьям у растений есть гениальная система проводимости, образованная очень специализированными клетками, которые в совокупности называются ксилемой.
Существует два основных типа проводящих трубок. Первый тип — трахеиды, представляющие собой очень узкие и удлиненные клетки, в которых прохождение веществ из одной клетки в другую осуществляется через окаймленные поры в клеточных стенках, расположенные на большой части этих стенок. Другим типом являются сосуды или трахеи с несколько большим диаметром и с законченными отверстиями (перфорациями) в зонах стенок, отделяющих от других клеток.
Хитрость заключается в использовании физических механизмов, которые не требуют вмешательства биологических процессов. Вода поднимается через трубки ксилемы благодаря нескольким физическим процессам, которые вместе образуют механизм, часто называемый натяжение-адгезия(прилипание)-когезия(сцепление). Основные идеи этого механизма были предложены более века тому назад Генри Диксоном. Механизм натяжение-адгезия-когезия основан на следующих явлениях или процессах.
Капиллярность
Капилярность — это любопытное явление, которое заставляет некоторые жидкости подниматься против гравитации спонтанно, внутри очень тонких протоков. Чем меньше диаметр канала, тем выше высота восходящего столба жидкости. Это движение, вызванное силой, возникшей в результате конфликта между двумя другими: поверхностное натяжение воды и адгезия воды к твердым поверхностям некоторых веществ:

- Когезия и поверхностное натяжение:
Молекулы воды, благодаря своей полярности, устанавливают между собой союзы, которые отвечают за сильное сцепление (объединение), существующее между ними.
Таким образом, сцепление заставляет слой молекул, контактирующих с воздухом, иметь натяжение, которое заставляет их вести себя как эластичная пленка.
Поверхность воды оказывает сопротивление деформации, поскольку ситуация максимального баланса будет такова, в которой поверхность контакта вода-воздух минимальна. Сила поверхностного натяжения отвечает за форму, которую принимают капли воды, а также за то, что легкие насекомые способны ходить по поверхности воды без погружения.
- Поверхностное натяжение и адгезия:
Между молекулами воды и твердыми поверхностями многих веществ также может быть установлена притягивающая сила, которая заставляет воду прилипать к этим поверхностям. Если адгезия к твердой поверхности сильнее когезии между молекулами воды, на краю жидкости, контактирующей с твердым материалом будет образовываться вогнутая кривизна поверхности.
Однако, поверхностное натяжение воды создает силу, которая тянет молекулы, образующие нижнюю часть кривой, чтобы попытаться уменьшить контактную поверхность воздух-вода, создавая плоскую пленку.
В результате создается возрастающая результирующая сила, которая приводит к небольшому подъему водного столба. Конечным результатом является то, что вода может буквально подняться по каналу, преодолевая силу тяжести.
Эвапотранспирация и натяжение
Капиллярность очень хорошо дает понять, как сырой сок может спонтанно подниматься через очень тонкие трубки ксилемы, но как только столб воды или сырой сок достигает конца трубки, капиллярности недостаточно, чтобы объяснить, как этот сок может продолжать подниматься из корня.
Механизм, который нам не ясен, связан с испарением воды, которая вырабатывается в листьях, и расходами этой воды при фотосинтезе.
Когда молекулы воды удаляются с конца трубки ксилемы из-за транспирации листьев, отверстие, как правило, немедленно заполняется другими молекулами, которые ниже. Это создает натяжение или отрицательное давление, которое буквально тянет столб воды вверх.
Сильное сцепление между молекулами воды предотвращает разрыв столбика сока внутри трубки, так что весь сок внутри трубки поднимается как единое целое. Естественно, из корней должна быть постоянная подача воды. В противном случае, как и в ситуациях засухи, столбик сока может быть прерван, что вызовет проблемы для растения.
Следующий рисунок объясняет явление натяжения, вызванного эвапотранспирацией:

Транспортировка переработанного сока через флоэму
К настоящему моменту мы не решили и более половины проблемы, потому что, как только фотосинтетические клетки образуют углеводы, необходимо распределить эти молекулы и их производные во многие другие отдаленные клетки, которые не способны их произвести.
Для этого распределения растения имеют другую ткань, называемую флоэмой, которая также образует каналы, проходящие через растение, но с некоторыми важными отличиями по сравнению с ксилемой.

Во-первых, проводящие клетки флоэмы живые, хотя они теряют свое ядро и довольно много органелл. В целом, они имеют меньший диаметр, чем проводники ксилемы.
Как и в ксилеме, клетки флоэмы выстраиваются рядами и соединяют свои стенки. В зонах соединения образуются структуры, называемые ситовидными пластинками. В этих областях имеются небольшие отверстия диаметром до 15 мкм (не такие большие, как перфорации ксилемы), которые позволяют передавать переработанный сок из одной ячейки в другую. Каналы, образованные объединением множества этих клеток, называются ситовидными трубками.
В течение некоторого времени считалось, что переработанные вещества могут транспортироваться путем диффузии через трубки флоэмы, но когда были сделаны расчеты, чтобы узнать скорость транспортировки через флоэму, стало ясно, что необходим другой механизм.
В настоящее время считается, что механизмом движения переработанного сока через флоэму является механизм, который предложил немецкий ученый Эрнст Мюнх в 1926 году. Этот механизм называется потоком под давлением и состоит в следующем.
Как только фотосинтетические клетки образуют органические молекулы, благодаря фотосинтезу, особенно глюкозу и фруктозу, большая часть этих молекул связывается с образованием сахарозы, которая является предпочтительной органической молекулой для переноса углеводов через растение.
Сахароза транспортируется во флоэму, из-за этого переработанный сок содержит высокую концентрацию этого вещества (сахароза составляет 90% веществ, присутствующих в переработанном соке, конечно, без учета воды). Поскольку концентрация сахарозы внутри флоэмы намного выше, чем снаружи, создается осмотическое давление, которое заставляет воду поступать во флоэму, тем самым увеличивая ее давление. Это давление подталкивает переработанный сок вдоль протоков флоэмы.
Резюме
Подводя итоги, можно отметить, что корни поглощают минеральные соли посредством активного транспорта, что генерирует осмотическое давление, вызывающее поглощение воды.
Эти вещества достигают проводников ксилемы, которые отвечают за транспортировку сырого сока от корней к фотосинтетическим структурам за счет механизма когезия-адгезия-натяжение.
После того как вырабатываются органические вещества, проводники флоэмы отвечают за транспортировку переработанного сока, богатого органическими молекулами, из листьев в другие области растения посредством механизма потока под давлением, основанного на механизмах осмотического давления.
Основным элементом ксилемы у высшего двудольного растения является сосуд.
Как и у ситовидных трубок, составляющие сосуд клетки-членики расположены наподобие звеньев цепи вдоль длинной оси органа, соединяясь друг с другом своеобразно модифицированными поперечными стенками. Членики сосудов во многих случаях также вытянуты, но не менее часто бывают и короткими.
Поперечные перегородки между члениками сосуда к тому времени, когда сосуд окончательно сформируется, продырявливаются, и протопласты клеток, формирующих сосуд, разрушаются. Если поперечные стенки члеников сосуда разрушаются полностью, образуется одно крупное отверстие, называемое простой перфорацией. Известны также и множественные перфорации, образующиеся в том случае, если оболочка не разрушается, а продырявливается во многих местах. Отверстия при множественной перфорации разбросаны в беспорядке (сетчатая перфорация) или располагаются правильными рядами (лестничная перфорация).
Утолщения на стенках сосудов могут быть весьма различного характера. Наиболее просто устроены спиральные утолщения, а также родственные им кольчатые утолщения. И те и другие свойственны наиболее рано возникающим анатомическим элементам ксилемы у высших растений. Древние формы растений с определенно выраженной ксилемой также имеют ксилемные элементы со спирально-кольчатыми утолщениями. От сосудов со спиральными утолщениями ряд постепенных переходов ведет к сосудам с округлыми окаймленными порами. На продольных разрезах через ксилему осевого органа какого-либо двудольного можно найти почти все градации утолщений. На рисунке при рассмотрении справа налево видны два крайних сосуда со спиральными утолщениями (4), причем более тонкий из них имеет несколько сильнее растянутые завороты спирали. Следующий сосуд (3) содержит так называемые лестничные утолщения. Сосуд (2) имеет вполне оформленные окаймленные поры— округлые и многочисленные. Это — пористый сосуд. Иногда переход от сосудов со спиральными утолщениями к пористым сосудам бывает очень резок (например, в стебле льна), чем нарушается онтогенетическая последовательность образования утолщений. Тип утолщений в значительной мере зависит от того, какой анатомический элемент находится по соседству с сосудом.
На рисунке изображен сосуд, к которому с одной стороны (3) примыкает паренхима, а с другой — либриформ (2), у которого вообще образуется мало пор.
Там, где к сосуду примыкает либриформ, на стенках сосуда пор совсем не образуется, а со стороны паренхимы стенки сосуда усеяны порами. Еще значительнее усложняется характер утолщений на стенках сосудов при соприкосновении их с клетками сердцевинных лучей, в свою очередь, достаточно разнообразных по очертаниям и структуре.
Кроме сосудов с продырявленными поперечными перегородками (перфорациями), в ксилеме большинства растений существуют другие водопроводящие элементы — трахеиды. Каждая трахеида — это отдельная мертвая прозенхимная клетка с более или менее заостренными концами. В отличие от сосудов поперечные стенки трахеид не разрушаются и перфораций не возникает. Утолщения стенок трахеид совершенно такие же, как и стенок сосудов. Среди трахеид встречаются клетки и со спиральными утолщениями, и с округлыми окаймленными порами. Трахеиды в некоторой мере обладают способностью к скользящему росту и могут врастать или между другими анатомическими элементами, или между другими трахеидами. Скользящим ростом обладают лишь молодые формирующиеся анатомические элементы.
Между трахеидами и либриформом существует ряд переходных форм. У семенных растений трахеиды в большем или меньшем количестве в зависимости от вида перемежаются с сосудами и другими элементами ксилемы. У папоротников, плаунов, хвощей и голосеменных растений трахеиды составляют почти всю массу ксилемы.
Подобно тому, как у ситовидных трубок имеются спутники, к сосудам часто примыкают паренхимные клетки, образующие обкладку сосуда. В зависимости от общей структуры ксилемы обкладка состоит или из одних паренхимных клеток, или в нее входят еще либриформ, трахеиды и более мелкие сосуды. Паренхима не только окружает сосуды, но и рассеяна среди ксилемных элементов; в последнем случае она называется древесинной паренхимой. Клетки ее имеют одревесневшие оболочки с простыми порами, но со стороны сосуда им соответствует окаймленная пора, которая сочетается с каждой простой порой. Протопласт клеток древесинной паренхимы очень долго не разрушается. Клетки паренхимы в ксилеме служат местом отложения запасов, так же как и паренхима, входящая в состав других тканей.
Иногда и на оболочках древесинной паренхимы развиваются окаймленные поры. Такие клетки не сохраняют протопласт в жизненном состоянии, так как служат передаточными пунктами в продвижении воды к соответствующим тканям.
Клетки паренхимы, непосредственно примыкающие к сосуду, находятся в тесном контакте с растворами, заполняющими сосуды, и в зависимости от условий то конденсируют в своих пластидах поступающие к ним углеводы и прочие вещества, то отдают эти вещества в полость сосуда. Специальными наблюдениями установлено, что в клетках паренхимы, примыкающих к сосуду и сообщающихся с ним через односторонние окаймленные поры, крахмал, сахар и прочие пластические вещества в течение года находятся в различных состояниях. Например, осенью и перед распусканием листьев крахмал в таких клетках находится в изобилии, а после распускания листьев содержание его значительно уменьшается. В клетках, не сообщающихся с сосудами при помощи пор, изменение состояния запасных веществ происходит очень медленно по сравнению с изменением их в клетках, сообщающихся с сосудами порами.
Вследствие того, что клетки древесинной паренхимы тесно связаны с сосудами и притом не только анатомически, но и физиологически, сосуды нельзя рассматривать совершенно изолированно. Это — система, в которой сосуд является только местом для продвижения весьма разведенных водных растворов. Предполагают, что состав растворов, а также отчасти направление их движения регулируются деятельностью паренхимных клеток, находящихся в непосредственном контакте с сосудам или входящих с ним в контакт при посредстве звеньев клеток. В сосудах многих растений, особенно древесных, сахар содержится не только весной, но и в другие времена года и иногда в заметных количествах (например, в условиях климата Закавказья — зимой). В зависимости от ряда условий клетки древесинной паренхимы, примыкающие к сосуду, могут образовывать выросты в полость сосудов — тиллы. Тиллы с течением времени разрастаются, заполняют всю полость сосуда, оболочки их одревесневают, и они становятся полным подобием создавшей их древесинной паренхимы, отличаясь от ее клеток лишь размерами и разнообразием очертаний. В таких тиллах откладываются те же запасные пластические вещества, что и в клетках древесинной паренхимы. Таким образом, сосуд с момента образования тилл тоже становится местом отложения запасных веществ.
Иногда тиллы весьма тесно заполняют полость сосуда, сильно сжимая друг друга и приобретая исключительно толстые оболочки, пронизанные поровыми каналами.
В этих случаях клетки тилл уже не похожи на паренхиму, расположенную вне полости сосуда. В некоторых редких случаях клетки тилл имеют облик и структуру типичных каменистых клеток. Тогда, плотно забивая сосуды и наполняясь различного рода отложениями, они придают ксилеме большую твердость. Особенно велико значение тилл в процессе формирования так называемого ядра древесины. Изредка тиллы образуются и у хвойных в трахеидах и смоляных ходах.
Клетки древесинной паренхимы в направлении, параллельном длинной оси органа, редко расположены беспорядочно, особенно клетки, не соприкасающиеся непосредственно с сосудом, с его стенкой. Такие клетки образуют подобие звеньев цепи, растянутой вдоль длинной оси органа. Но вертикальные ряды, составленные из клеток древесинной паренхимы, — короткие, оканчивающиеся с обеих концов клетками с заостренными окончаниями, подобно трахеидам.
Древесинная паренхима многолетних растений по своему расположению в толще годичного слоя подразделяется на два типа: паратрахеальную, приуроченную к сосудам, и апотрахеальную, не связанную с сосудами. В пределах паратрахеального типа различают вазицентрическую, крыловидную и сомкнуто-крыловидную паренхиму. Эти термины имеют чисто описательное значение и определяют характер расположения клеток вокруг сосудов.
В пределах апотрахеального типа по расположению составляющих элементов различают диффузную, метатрахеальную и терминальную паренхимы. Диффузная паренхима на поперечных срезах обнаруживается в виде одиночных клеток, рассеянных среди клеток лучей. В продольном направлении клетки диффузной паренхимы располагаются в виде цепочек. В том случае, если несколько таких цепочек располагаются рядом, древесинная паренхима называется метатрахеальной. На поперечных срезах она имеет вид тангентальных полосок, состоящих из одного ряда клеток. У многих древесных растений с сезонным приростом древесинная паренхима развивается только к концу прироста. При этом она формируется в виде более или менее сплошного слоя вокруг кольца прироста. Такая паренхима называется терминальной.
Клетки древесинной паренхимы образуются так называемыми веретенообразными клетками камбия. Если клетка камбия непосредственно дифференцируется в паренхимную клетку, последняя имеет веретеновидную форму, напоминая элементы либриформа. Такие клетки носят название заменяющих волокон и отличаются от либриформа простыми округлыми порами и остающимся в жизнедеятельном состоянии протопластом.
Однако чаще перед образованием паренхимы клетка камбия несколько раз делится в одном направлении; возникшая цепочка клеток дифференцируется как тяжевая паренхима. Сопоставляя анатомические элементы флоэмы с анатомическими элементами ксилемы, можно убедиться, что и флоэма и ксилема составлены из элементов трех основных типов, приспособленных к выполнению соответствующих функций: запасающей, проводящей и механической. Между этими тремя основными типами существуют переходные как в структурном, так и в функциональном отношении. Так, анатомическим элементом, предназначенным преимущественно для отложения и хранения различных запасных веществ, является паренхима, лубяная во флоэме и древесинная в ксилеме. Очертания паренхимных клеток в обоих участках проводящей системы разнообразны; причем различие заключается в особенностях строения оболочек. Клетки лубяной паренхимы имеют целлюлозные оболочки, а клетки древесинной паренхимы — одревесневшие. Поры в обоих типах паренхимы простые, если протопласт клетки находится в жизнедеятельном состоянии. В клетках древесинной паренхимы с отмершим протопластом поры могут быть и окаймленными, причем степень развития и рисунок окаймленных пор у таких клеток различны.
Механическая система как во флоэме, так и в ксилеме представлена волокнами. У некоторых растений в коре встречаются паренхимные толстостенные клетки (каменистые), но в самой флоэме таких клеток не образуется; они характерны для коровых участков сердцевинных лучей.
Между лубяными волокнами флоэмы и либриформом структурная аналогия еще более полная, чем между анатомическими элементами запасающей и проводящей систем. Оболочки лубяных волокон также нередко одревесневают, уподобляясь в этом отношении либриформу. Поры у таких лубяных волокон щелевидные и косые (вариант простых пор), вполне аналогичные порам клеток либриформа.
Оболочки механических элементов флоэмы и ксилемы толстостенные. У некоторых растений как лубяные волокна, так и волокна либриформа имеют поперечные перегородки.
Вообще анатомические элементы флоэмы и ксилемы, подобно веем элементам растительных тканей, очень пластичны и представлены рядом переходных форм. Особенно много переходных
форм наблюдается в узлах стеблей — в тех местах, где от стебля отходят листья и ветки, а также в местах поранения, где происходит заживление ран.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
В растительном организме, так же как и в организме животных имеется транспортные системы, обеспечивающие доставку питательных веществ по назначению. На сегодняшнем занятии разговор пойдёт о проводящих тканях растения.
Проводящие ткани – ткани, по которым происходит массовое передвижение веществ, возникли как неизбежное следствие приспособление к жизни на суше. От корня к листьям движется восходящий, или транспирационный, ток водных растворов солей. Ассимиляционный, нисходящий ток органических веществ направляется от листьев к корням. Восходящий ток осуществляется почти исключительно по сосудам древесины (ксилемы), а нисходящий – по ситовидным элементам луба (флоэмы).

1. Восходящий ток веществ по сосудам ксилемы 2. Нисходящий ток веществ по ситовидным трубкам флоэмы

Клетки проводящей ткани характеризуются тем, что они вытянуты в длину и имеют форму трубочек с более или менее широким диаметром (в общем, напоминают сосуды у животных).
Существуют первичные и вторичные проводящие ткани.
Вспомним классификацию тканей на группы по форме клеток.

Ксилема и флоэма – это сложные ткани, состоящие из трёх основных элементов.

Проводящие элементы ксилемы.
Наиболее древними проводящими элементами ксилемы являются трахеиды (рис.1)– это вытянутые клетки с заостренными концами. Они дали начало древесинным волокнам.
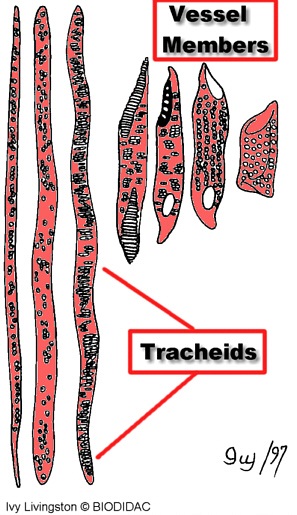
Трахеиды имеют одревесневшую клеточную стенку с различной степенью утолщения, кольчатую, спиралевидную, точечную, пористую и т.д. форму (рис. 2). Фильтрация растворов происходит через поры, поэтому передвижение воды в системе трахеид совершается медленно.
Трахеиды встречаются у спорофитов всех высших растений, а у большинства хвощевидных, плауновидных, папоротниковидных и голосеменных, являются существенными проводящими элементами ксилемы. Прочные стенки трахеид позволяют им выполнять не только водопроводящие функции, но и механические. Часто они являются единственными элементами, придающими органу прочность. Так, например, у хвойных деревьев в древесине отсутствует специальная механическая ткань, и механическая прочность обеспечивается трахеидами.
Длина трахеид колеблется от десятых долей миллиметра до нескольких сантиметров.

Рис. 2 Трахеиды и их расположение относительно друг друга
Рис. 2 Трахеиды и их расположение относительно друг друга
Рис. 3 Расположение первичных и вторичных проводящих тканей в корне
Расположение первичных и вторичных проводящих тканей в стебле
Первые по времени образования сосуды (рис. 3) – протоксилема – закладываются на верхушке осевых органов, непосредственно под верхушечной меристемой, там, где окружающие их клетки ещё продолжают вытягиваться. Зрелые сосуды протоксилемы способны растягиваться одновременно с вытягиванием окружающих клеток, поскольку их целлюлозные стенки ещё не сплошь одревеснели – лигнин (особое органическое вещество, вызывающее одревесневание стенок клеток) откладывается в них кольцами или по спирали. Эти отложения лигнина позволяют трубкам сохранять достаточную прочность во время роста стебля или корня.
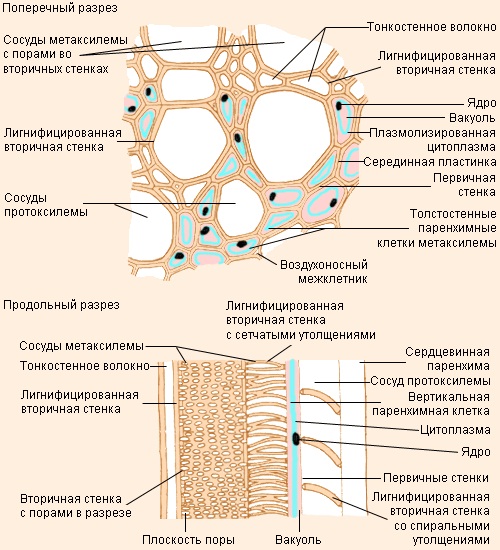
Рис. 4 утолщения клеточных стенок сосудов
С ростом органа появляются новые сосуды ксилемы, которые претерпевают более интенсивную лигнификацию и завершают своё развитие в зрелых частях органа, — формируется метаксилема. Тем временем самые первые сосуды протоксилемы растягиваются, а затем разрушаются. Зрелые сосуды метаксилемы не способны растягиваться и расти. Это мёртвые, жёсткие, полностью одревесневшие трубки. Если бы их развитие завершилось до того, как закончилось вытягивание окружающих живых клеток, то они бы очень сильно мешали этому процессу.
Утолщения клеточных стенок сосудов так же, как и у трахеид, бывают кольчатыми, спиральными, лестничными, сетчатыми и пористыми (рис. 4 и рис. 5).
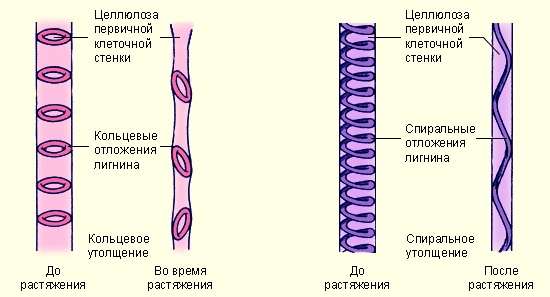
Рис. 5 Типы перфорации сосудов
Длинные полые трубки ксилемы – идеальная система для поведения воды на большие расстояния с минимальными помехами. Так же как и в трахеидах, вода может переходить из сосуда в сосуд через поры или через неодревесневающие части клеточной стенки. Вследствие одревесневания клеточные стенки сосудов обладают высокой прочностью на разрыв, что тоже очень важно, потому что благодаря этому трубки не спадаются, когда вода движется в них под натяжением. Вторую свою функцию – механическую – ксилема также выполняет благодаря тому, что она состоит из ряда одревесневших трубок.
Проводящие элементы флоэмы. Ситовидные трубки образуются из прокамбия в первичной флоэме ( протофлоэма) и из камбия во вторичной флоэме ( метафлоэма). По мере того как растут окружающие её ткани, протофлоэма растягивается и значительная её часть отмирает, перестает функционировать. Метафлоэма созревает уже после того, как закончится растяжение.
Членики ситовидных трубок имеют весьма характерное строении. У них более тонкие клеточные стенки, состоящие из целлюлозы и пектиновых веществ, и этим они напоминают паренхимные клетки, однако их ядра при созревании отмирают, а от цитоплазмы остаётся только тонкий слой, прижатый к клеточной стенке. Несмотря на отсутствие ядра, членики ситовидных трубок остаются живыми, но их существование зависит от примыкающих к ним клеток-спутниц, развивающихся из одной с ними меристематической клетки (рис. 6).
Вопрос: — Какие клетки животных, являясь безъядерными, также остаются живыми?
Членик ситовидной трубки и его клетка-спутница составляют вместе одну функциональную единицу; у клетки-спутницы цитоплазма очень густая и отличается высокой активностью, на что указывает присутствие многочисленных митохондрий и рибосом. В структурном и функциональном отношении клетка-спутница и ситовидная трубка тесно связаны и совершенно необходимы для их функционирования: в случае гибели клеток-спутников погибают и ситовидные элементы.

Рис. 6 Ситовидная трубка и клетка спутница
Характерной чертой ситовидных трубок является наличие ситовидных пластинок (рис. 7). Эта их особенность сразу же бросается в глаза при рассматривании в световом микроскопе. Ситовидная пластинка возникает на месте соединения торцевых стенок двух соседних члеников ситовидных трубок. Вначале через клеточные стенки проходят плазмодесмы, но затем их каналы расширяются и образуют поры, так что торцевые стенки приобретают вид сита, через которое раствор перетекает из одного членика в другой. В ситовидной трубке ситовидные пластинки располагаются через определённые промежутки, соответствующие отдельным членикам этой трубки.
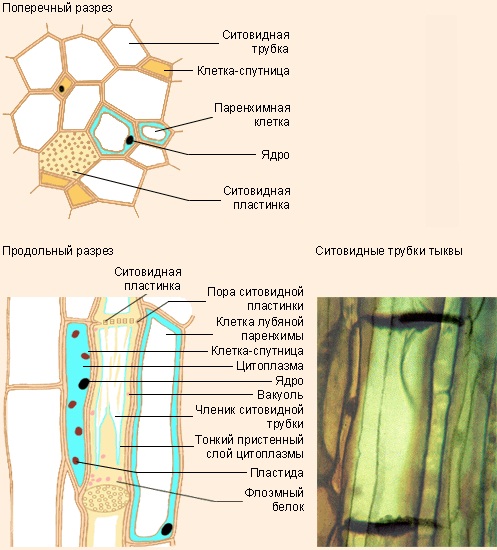
Рис. 7 Ситовидные пластинки ситовидных трубок
Основные понятия: Флоэма (протофлоэма, метафлоэма), ситовидные трубки, клетки-спутницы. Ксилема (протоксилема, метаксилема) трахеиды, сосуды.
Читайте также: