
В чем заключается прогрессивные черты строения покрытосеменных
Обновлено: 07.07.2024
Общая характеристика. Покрытосеменные, или Цветковые растения, – это крупнейший отдел современных высших растений, насчитывающий около 250 тыс. видов. Однако все разнообразие этого отдела человечеству еще не известно: каждый месяц в мире описывают более 100 новых видов покрытосеменных растений. В настоящее время покрытосеменные играют решающую роль в формировании растительного покрова Земли. Они растут во всех климатических зонах и входят в состав практически всех биогеоценозов земного шара. Это свидетельствует об их высокой приспособленности к современным условиям существования на Земле.
Появившись в меловом периоде (в конце мезозойской эры), в начале кайнозоя покрытосеменные растения уже господствовали на всем земном шаре, резко потеснив папоротники и голосеменные растения. Рассмотрим, какие же особенности строения и развития позволили покрытосеменным занять господствующее положение на Земле.
Прогрессивные черты организации покрытосеменных растений.
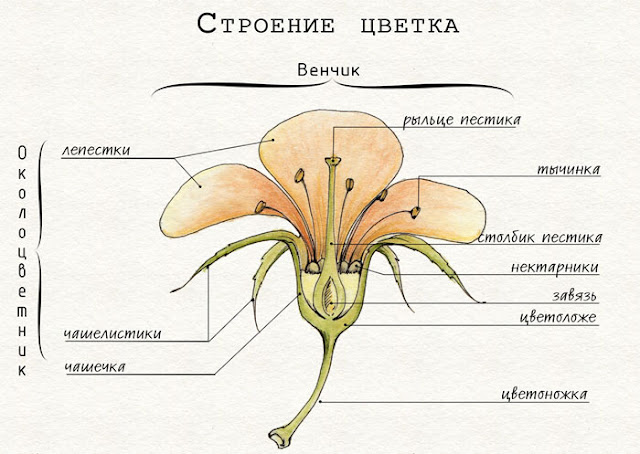
Появление цветка. Главной и уникальной особенностью покрытосеменных является наличие цветка. Огромное разнообразие цветков не дает возможности свести все существующие сейчас типы цветков к единой исходной предковой модели. Возможно, в процессе эволюции формирование цветков разных видов происходило параллельно различными путями. Существует несколько гипотез происхождения цветка, одна из которых в настоящее время является ведущей. Согласно этой гипотезе, цветки, как и стробилы (шишки) голосеменных растений, возникли из спороносных побегов древних семенных папоротников. В дальнейшем эволюция шишек голосеменных и цветков покрытосеменных растений происходила параллельно независимо друг от друга. Итак, цветок – это видоизмененный укороченный спороносный побег, все части которого, кроме цветоложа, имеют листовую природу. Мегаспорофиллы превратились в плодолистики, которые, срастаясь между собой, сформировали пестик (пестики), микроспорофиллы преобразовались в тычинки, а окружающие их стерильные листья образовали околоцветник. Цветок – это орган размножения (бесполого и полового), в котором возникают микро- и мегаспоры, формируются заростки, происходит опыление и оплодотворение, развивается зародыш спорофита и запасающая ткань, и в итоге образуются семя и плод.
Наличие завязи. Семязачатки покрытосеменных развиваются во влажной камере – завязи. Это предохраняет их от неблагоприятных воздействий среды, в первую очередь от высыхания, что очень важно в условиях жизни на суше. Из завязи образуется плод, внутри которого заключены семена. Расположение семян внутри плода определило название отдела – Покрытосеменные. Плоды обеспечивают защиту семян и способствуют их распространению.
Редукция гаметофитов. Покрытосеменным свойственна дальнейшая редукция мужского и женского гаметофитов. Заростки не имеют не только вегетативных частей, но и половых органов – гаметангиев. Мужской гаметофит цветковых растений – это пыльцевое зерно, а женский гаметофит представлен зародышевым мешком.
Двойное оплодотворение. У покрытосеменных растений в оплодотворении участвуют два спермия, т. е. осуществляется двойное оплодотворение. В результате образуются диплоидный зародыш и триплоидный эндосперм – питательная ткань для зародыша.
Развитие проводящих тканей. У цветковых растений прекрасно развита проводящая система. В древесине (ксилеме) транспорт воды и растворенных в ней минеральных веществ осуществляется преимущественно по сосудам, а не по трахеидам, как у голосеменных растений. Проводящие элементы флоэмы представлены уже не ситовидными клетками, а ситовидными трубками, рядом с которыми располагаются клетки-спутницы (сопровождающие клетки).
Разнообразие жизненных форм. В отличие от голосеменных, которые представлены только древесными растениями, жизненные формы покрытосеменных растений значительно разнообразнее. Современные ученые считают, что первые цветковые были древесными растениями. Из них возникли кустарники и кустарнички. В дальнейшем, обладая наиболее совершенной системой размножения, покрытосеменные растения расселились по всей планете. Попадая в различные условия, они были вынуждены приспосабливаться, чтобы выжить. В засушливых условиях, за полярными широтами и на высокогорье преимущество получали растения, успевавшие сформировать вегетативные органы, отцвести и образовать семена за один вегетационный период. Поэтому одной из наиболее важных форм адаптации стало появление травянистых растений. В зависимости от продолжительности жизни травянистые растения подразделяют на однолетние, двулетние и многолетние. Однолетние отмирают после образования плодов. Двулетние в первый год жизни формируют вегетативные органы, запасают питательные вещества (как правило, в подземных видоизмененных побегах или корнях), а на второй год цветут и плодоносят. Многолетние травы живут и плодоносят в течение нескольких лет, но каждый год по окончании вегетационного периода их надземные побеги отмирают, а подземные переживают неблагоприятные условия, чтобы на следующий год дать начало новым надземным побегам. Считают, что травы возникли в результате значительного ослабления деятельности камбия, что привело к постепенному сокращению численности древесных растений.
Огромное морфологическое разнообразие листьев, корней, побегов покрытосеменных растений было рассмотрено нами ранее при изучении строения их вегетативных органов. Многообразие цветков мы подробно рассмотрим позже.
Прогрессивные изменения структуры тканей, вегетативных и генеративных органов обеспечили возникновение огромного видового разнообразия. Высокая эволюционная пластичность позволила цветковым растениям распространиться по всему земному шару и приспособиться к самым разным экологическим условиям.
Покрытосеменные – это единственная группа растений, образующая сложные многоярусные сообщества, что способствует более интенсивному использованию всех условий среды и более успешному освоению новых территорий.
Значение покрытосеменных растений. Значение покрытосеменных растений в природе и жизни человека столь велико, что почти невозможно перечислить все те отрасли народного хозяйства, где они используются в том или ином виде. Пищевые и декоративные, лекарственные и кормовые, дающие строительный материал и сырье для химической промышленности – цветковые растения обеспечили развитие и существование современных цивилизаций. Мы рассмотрим более подробно практическое значение покрытосеменных, изучая отдельные семейства.
Однако роль растений в жизни человека гораздо шире, нежели только практическое использование. Покрытосеменные растения определяют состояние современной биосферы. Они образуют основную биомассу земного шара, насыщают атмосферу кислородом, формируют почву. Поглощая и испаряя в атмосферу огромное количество воды, участвуют в формировании климата. Служат основой большинства биогеоценозов.
Предки покрытосеменных, как указывалось ранее, остаются до настоящего времени неизвестными. Но, несомненно, вся эта группа возникла от голосеменных и обособилась от них, вероятно, еще на ранних этапах всей истории семенных растений. Из современных покрытосеменных, соответственно стробилярной теории цветка, по-видимому, наиболее близок к древнейшим покрытосеменным подкласс Magnoludae .Однако Magnolndae имеют уже настолько специализированный аппарат для привлечения насекомых-опылителей, что естественно предположить существование в прошлом еще более древней группы покрытосеменных. Такие гипотетические предковые покрытосеменные могли быть неустойчиво энтомофильными растениями, у которых перенос пыльцы совершался и ветром ,насекомыми. По всей вероятности, у них достаточно широко было распространено и самоопыление.
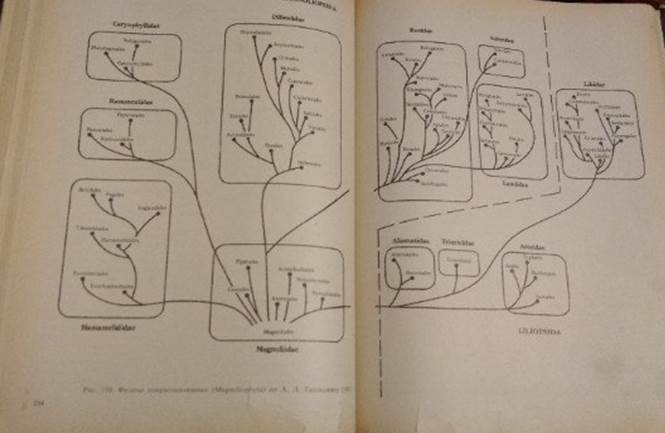
Схема эволюции цветковых растений
Гипотетические покрытосеменные предки современных цветковых дали начало, вероятно, нескольким линиям эволюции . Главной из них, т. е. имевшей впоследствии наибольший успех в освоении самых разнообразных условий жизни, была линия, в которой рано выработались Magnoliidae. Другой была линия, представленная подклассом Hamamelidae (см.рис.)
В конце прошлого — начале текущего столетия было широко распространено представление об однопокровных, которые в основном соответствуют Hamamelidae, как о группе весьма древней, примитивной, близкой к предкам покрытосеменных.Учение о происхождении покрытосеменных от голосеменных через однопокровные дополнялось специально разработанной австрийским ботаником Р.Веттштейном теорией происхождения цветка покрытосеменных из "соцветия" голосеменных (типа эфедры). Эта теория получила название теории псевданция (ложного цветка).
Научная критика концепции происхождения всего многообразия покрытосеменных от однопокровных вскрыла прежде всего, что морфологически "простое" строение цветка может быть следствием упрощения, возникшего в связи с особым направлением отбора. Это подтверждается наличием остаточного околоцветника (дуб, лещина), появлением в цветках рудиментов недостающего пола и развитием в качестве исключения обоеполых цветков вместо однополых.
Если простота цветков однопокровных вторичная, то приходится заключить, что эта группа прошла длительный путь эволюции, возникнув от предков с обоеполыми цветками. Но обоеполость цветков тесно связана с энтомофилией. Поэтому и распространенная у однопокровных анемофилия должна была прийти на смену энтомофилии, что возможно в процессе приспособления к жизни большими одновидовыми скоплениями.По-видимому, именно благодаря этому от каких-то энтомофильных лилиецветных возникли анемофильные осоки и злаки; анемофильные полыни, несомненно, являются производными энтомофильных сложноцветных.
Подвергалась глубокой критике и псевданциевая теория происхождения цветка, оказавшаяся неубедительной.В итоге, на смену представлению о первичности и примитивности однопокровных сформировалось прямо противоположное воззрение — однопокровных стали рассматривать как группу, прошедшую длительный путь эволюции и возникшую от высокоспециализированных покрытосеменных.При этом допускалось, что различные порядки однопокровных — производные разных эволюционных линий покрытосеменных. Это означало отрицание группы как единого естественного таксона.Схематический обзор родственных связей покрытосеменных растений позволяет сделать несколько обобщающих выводов. Ход эволюционного развития осуществлялся в виде нескольких параллельных потоков, которые соответствуют подклассам и классам.
Таким образом, система связей между покрытосеменными не может быть уподоблена родословному дереву; она, скорее, соответствует "кустарнику" на том этапе его развития, когда он уже потерял главный корень. Но в таком случае приходится признавать, что однокоренное, монофилетическое происхождение покрытосеменных продолжает оставаться гипотезой, хотя и очень вероятной. Гипотетичность монофилетического происхождения покрытосеменных усугубляется и тем, что пока нет возможности безоговорочно указать группу древних растений, вероятнее всего голосеменных, которая дала начало первым цветковым. При таких обстоятельствах не приходится удивляться, что по временам возникали и продолжают появляться гипотезы о полифилетическом возникновении цветковых. Однако полифилетические воззрения находятся не в лучшем положении, так как выдвигают задачу отыскания не одного, а нескольких предков покрытосеменных, что не смягчает, а обостряет трудность проблемы.
В схеме родственных отношений покрытосеменных растений достаточно четко отражен параллелизм в развитии подклассов и даже классов. Одно из его проявлений состоит в том, что почти в каждом подклассе имеются энтомофильные и анемофильные ветви. Распространенность анемофильных приспособлений в разных эволюционных потоках свидетельствует о том, что существуют какие-то общие условия, при которых анемофилия представляет существенное преимущество в борьбе за существование. Одно из таких условий — способность произрастания массовыми одновидовыми скоплениями, в которых опыление ветром, при производстве большого количества пыльцы, перестает быть ненадежным. Но произрастание в форме массовых скоплений возможно при условии,что внутривидовая конкуренция становится инструментом саморегуляции численности, поддерживающим ее на уровне, необходимом для подавления конкурентов. С этой точки зрения устойчивое ветроопыление, если оно обеспечивается надежными микробиологическими особенностями цветков и соцветий, представляет высокое достижение в ходе исторического развития цветковых.Но все же более распространенным у покрытосеменных оказалось опыление животными: насекомыми, птицами, рукокрылыми.
Почти в каждом подклассе можно найти растения (главным образом зоофильные) с правильными и неправильными (зигоморфными) цветками, с раздельнолепестным венчиком, с верхней и нижней завязью. На основании морфогенеза, спайнолепестности, зигоморфности и нижней завязи можно с уверенностью сказать, что эти признаки являются более поздними, вторичными по сравнению с первичными: раздельнолепестностью и актиноморфностью цветка, верхней завязью. Эти данные, принимая во внимание приспособительное значение вторичных признаков, помогают филогенетику размещать порядки и семейства в пределах эволюционных рядов соответственно формированию во времени. Однако факт, что указанные вторичные морфобиологические особенности имеют место в энтомофильной эволюции во всех подклассах, приходится рассматривать как закономерность. Нет необходимости видеть причину этого во "внутренних свойствах" растений, "предопределяющих", как иногда думают, эволюционные изменения. Возможности изменений органов у растений в целом очень ограничены.
Сдвиги в онтогенезе цветка в сторону более раннего наступления базального роста у оснований примордиев лепестков, что приводит к возникновению спайнолепестности, — достаточная основа для последующего отбора, поскольку трубка венчика ограничивает круг потребителей продуктов, вырабатываемых цветком, и одновременно защищает эти продукты от посетителей, которые не могут произвести опыление. Аналогичное можно сказать и о возникновении других вторичных особенностей цветка.
Основные контуры эволюционных рядов покрытосеменных растений были построены главным образом на основе учета разнообразия цветков и плодов. В настоящее время эти ряды или те, которые получены в результате критического пересмотра ранее построенных, все чаще находят подтверждение в особенностях вегетативного тела. В этом отношении очень важные результаты были получены при анатомическом изучении покрытосеменных растений, в частности их водопроводящей системы. .
Настоящие сосуды, как известно, свойственны только покрытосеменным. Если подобные образования и встречаются в более древних группах (голосеменные, папоротникообразные), то это только аналоги сосудов. Но членики сосудов покрытосеменных различны. Среди них можно отличать и более типичные, и приближающиеся к трахеидам. Кроме того, известно, что среди покрытосеменных есть и бессосудистые растения. Такие группы естественно рассматривать как более древние. В некоторых эволюционных рядах, отвечающих прогрессивному изменению цветка, обнаруживается последовательное нарастание признаков совершенствования водопроводящей аппаратуры растения. Подтверждаются эти ряды и другими анатомическими признаками.
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
В течение последних десятилетий большого прогресса достигла особая ветвь морфологии — морфология пыльцы. Ее данные, привлекаемые к построению филогенетических рядов, помогают ориентироваться в высоте организации, а также в наличии сходств, которые можно истолковывать как показатели родства.Хотя представления о ходе эволюционного развития, показанные прилагаемым схематическим рисунком, нельзя считать окончательным, все же в основном они правильно отражают, во-первых, взаимосвязь по крайней мере более крупных подразделений покрытосеменных, а во-вторых, размещение в каждом из стволов более поздних (по возникновению в пределах данной филогенетической ветви) и более ранних таксонов.
Эта схема позволяет выделить в многообразии покрытосеменных три уровня. Первый, наиболее низкий уровень включает примитивных покрытосеменных, их неизвестных предков и обширный подкласс Magnoliidae. Этому уровню противоположен другой, наивысший, характеризуемый спайнолепестностью и часто зигоморфностью цветка, нижней завязью, тенденцией к мельчанию цветков и образованию компактных соцветий. Этот уровень достаточно ясен у энтомо- фильных двудольных. У высших энтомофильных однодольных соответственно наблюдается зигоморфность цветков, сопровождаемая порой глубокими перестройками андроцея и гинецея, а также появление сложных аппаратов, обеспечивающих анемофилию. Между этими двумя уровнями находит себе место промежуточный, в котором отразилось много частных проявлений эволюционного процесса.
Охватывающая эволюцию растений по геологическим периодам
Домашние и дикие растения, их описание и выращивание; болезни и вредители. Интересные сведения из ботаники и других областей науки. Доклады для школьников. Природа Приморского края (растения и животные). История, культура, достопримечательности Владивостока и России в целом.
Вечерний Владивосток / улица Адмирала Фокина

- Главная страница
- Всё о бегониях
- Всё о бегониях - 2
- Список бегоний - 1
- Список бегоний - 2
- Список бегоний - 3
- Список бегоний - 4
- Список бегоний - 5
- Список бегоний - 6
- Список бегоний - 7
- Список бегоний - 8
- Список бегоний - 9
- Мои растения
- Доклады
- В помощь цветоводам, садоводам и огородникам
- Опознавалка бегоний и других растений
- Дизайн в саду и дома
- Комнатные растения
- Садовые растения
- Растения по алфавиту
- Растения по алфавиту (2)
- Приморский край
- Приморский край - 2
- Латынь в ботанике
- Биология
- Ботаники
- История и культура
- Интересное
- Вопрос-ответ
воскресенье, 7 июня 2015 г.
Покрытосеменные (Angiospermatophyta)
Представляют собой самый крупный отдел растительного мира. Насчитывают более 250 тысяч видов, объединённых в 13 тысяч родов и 533 семейства.
Возникнув приблизительно 130 млн. лет назад, эти растения быстро распространились по всем континентам и в настоящее время на большей части суши играют решающую роль в формировании растительного покрова. Произрастают покрытосеменные во всех климатических зонах и в различных экологических условиях: в тропических лесах и в тундре, на болотах и в пустынях, в прибрежных зонах морей и в горах. У них максимально развита стратегия патиентности ( выносливость к экстремальным условиям ) и эксплерентности ( способности к быстрому заселению не занятых местообитаний и восстановлению нарушенной растительности ). Такие огромные потенциальные возможности покрытосеменных связаны с целым рядом прогрессивных изменений в их структуре и организации.
В отличие от голосеменных, цветковые представлены большим разнообразием жизненных форм и способны формировать многоярусные растительные сообщества . Среди них есть деревья, кустарники, полукустарники; многолетние, двулетние и однолетние травы; лианы, поднимающиеся в полог и растения-эпифиты. Есть паразиты, и даже сапрофиты.

В отличие от предшествующих им групп растений, покрытосеменные имеют разнообразные типы корневых систем (стержневую, мочковатую, смешанную, ветвистую) и способны формировать помимо земляных водяные, воздушные корни и корни–присоски (гаустории). Вступая в симбиоз с грибами микоризообразователями, бактериями ризосферы, а иногда и азотофиксирующими бактериями , корни цветковых обеспечивают максимально эффективное поглощение воды и минеральных веществ. Стебли покрытосеменных характеризуются большим разнообразием форм ветвления и способов роста (прямостоячие, ползучие, цепляющиеся и т.д.). Разнообразны и формы простых и сложных листьев, их расположение, жилкование, расчленение. Таким образом, стебли и листья этих растений создают обширную ассимиляционную поверхность, адаптированную к различной интенсивности освещения и позволяющую максимально эффективно использовать энергию солнечного света .
Листопадность – свойство, которое характерно лишь некоторым голосеменным, позволяет большинству покрытосеменных переносить засушливый период жаркого климата и холод суровых зим умеренной климатической зоны. Кроме того, корни и побеги этих растений способны видоизменяться и выполнять необычные функции : защитную, запасающую, ловли насекомых и т.д. У цветковых хорошо развито вегетативное размножение . А размножение при помощи клубней, луковиц, корневищ способствует не только увеличению числа особей, но и сохранению вида при воздействии неблагоприятных факторов внешней среды (заморозков, пожаров и т.д.).

Перестройка коснулась и внутреннего строения вегетативных органов. Так, у покрытосеменных появились более совершенные проводящие ткани : трахеи (сосуды) и ситовидные трубки с клетками-спутницами, обеспечивающие эффективный транспорт веществ, как по восходящему, так и по нисходящему токам. Новые виды механических тканей – колленхима, древесные и лубяные волокна – придают прочность не только древесным, но и травянистым растениям. Совершенствуется внутренняя структура листа : мезофилл листа дифференцирован на столбчатую и губчатую паренхиму. Лист пронизан целой сетью сосудисто-волокнистых проводящих пучков, эффективно обеспечивающих снабжение листьев водой и отток из них, синтезированных органических веществ. Наконец, для цветковых характерно накопление в клетках тканей различных химических веществ (алкалоидов, гликозидов, сапонинов, эфирных масел и др.), которые обеспечивают им биохимическую защиту от большинства фитофагов.


Для цветковых характерны разнообразные приспособления к различным способам перекрёстного опыления и ограничению самоопыления : двудомность, самонесовместимость, обусловленная ингибированием, т. е. задержкой прорастания собственной пыльцы, херкогамия (пространственное разобщение пыльников и рыльца), дихогамия (неодновременное созревание тычинок и пестиков: протерандрия - раньше созревают тычинки, протерогиния – раньше созревают пестики). При этом у многих видов самоопыление сохраняется как запасной вариант на случай, если перекрестное опыление не произошло. Есть среди цветковых и исключительно самоопыляемые растения (арахис).
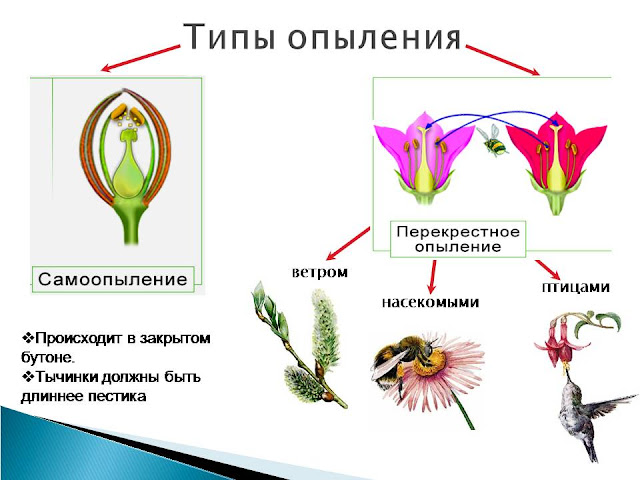
Только для цветковых характерно двойное оплодотворение , биологический смысл которого заключается в том, что развитие триплоидной питательной ткани – вторичного эндосперма происходит лишь в том случае, если произойдёт оплодотворение. Нет зародыша – нет напрасных расходов внутренних ресурсов растения на формирование многоклеточной, питательной ткани. Возможно отложение запасных питательных веществ и в семядольных листьях зародыша, и в ткани мегаспорангия (перисперме).

У представителей некоторых семейств (например, Астровых) распространено явление апомиксиса – развитие семени без оплодотворения, а у других явление многозародышевости - в одном семени формируется несколько зародышей (цитрусовые, клевера). Наконец, семена у большинства цветковых способны переносить неблагоприятные условия, находясь, длительное время в состоянии покоя.

Систематика покрытосеменных является частью общего раздела ботаники – систематики растений , зарождение которой относят ко времени расцвета древнегреческой культуры. Таким образом, история её развития насчитывает более двух тысячелетий и включает в себя периоды создания утилитарных, искусственных и естественных систем. Необходимость создания филогенетической системы цветковых растений, отражающей не только родство определенных систематических групп, но и пути их исторического развития, была очевидна давно, а в настоящее время является одной из основных задач современной биологии. Такая система необходима для успешного развития новых биологических дисциплин (биохимии, молекулярной биологии), т. к. она даёт ясную картину филетических взаимоотношений между различными формами жизни; для решения проблем современной экологии, которая при изучении экосистем опирается на таксономический состав и эволюционные взаимоотношения их компонентов; для охраны биоты и поиска новых растений, пригодных для сельского хозяйства, селекции, техники, медицины и пр.
Можно привести много примеров того, как на основе таксономической близости видов удалось предположить и обнаружить в них сходные химические соединения. Так, сравнительный биохимический анализ двух растений из семейства Валериановые – валерианы аптечной ( Valeriana officinales ), природные запасы которой незначительны, и довольно часто встречающейся патринии скабиозолистной ( Patrinia skabiosifolia ) показал, что патриния во многом сходна по содержанию действующих веществ и свойствам с традиционно используемой валерианой. Таким образом, был найден новый источник лекарственного сырья.
Сложности создания филогенетической системы наиболее ярко проявились в 60-70 годы, когда лавинообразно обрушился поток информации, полученной в результате применения новых методов исследования растений – кариологического, анатомического, субмикроскопического, биохимического, палинологического. Тем не менее, к концу 20 века был достигнут значительный успех в создании макросистемы покрытосеменных. Большинство современных систематиков основываются на эвантовой (стробилярной) теории происхождения цветка, согласно которой, цветок представляет собой видоизменённый укороченный спороносный побег, или обоеполый стробил голосеменных. Исходя из этого принципа, составлен кодекс признаков примитивной и прогрессивной организации цветковых растений, отражающий особенности строения не только генеративных, но и вегетативных органов растений (табл.1).
В настоящее время покрытосеменные, или цветковые, растения преобладают среди растительности на Земле. Их насчитывается около 300 тыс. видов. Покрытосеменные — наиболее высокоорганизованные растения. Они представлены разными жизненными формами (деревьями, кустарниками, травами и др.) с разной продолжительностью жизни и заселяют большинство сухопутных мест обитания.
Покрытосеменные, или цветковые, растения — это группа, находящаяся в статусе отдела по современной классификации.
От всех других растений покрытосеменные отличаются наличием у них:
более совершенным строение проводящей системы.
Кроме этих основных, есть и другие отличительные признаки.
У покрытосеменных растений образуются семена, в отличие от мхов и папоротников, у которых образуются споры. Однако семена образуются не только у покрытосеменных, но также у голосеменных растений. Главное отличие между ними заключается в отсутствии у голосеменных завязи, защищающей семязачатки, и плода, служащего органом распространения семян.
Приобретение в процессе эволюции цветка и плода позволило покрытосеменным (цветковым) освоить новые способы опыления генеративных органов и новые способы распространения семян. Опыление у них стало осуществляться не только с помощью воды (как у мхов и папоротников) и ветра (как у голосеменных). Преимущественно покрытосеменные приспособились к опылению разнообразными насекомыми. Для этого у них эволюционно развились яркие, пахнущие, вырабатывающие нектар хорошо заметные цветки и соцветия. Наличие плода, в свою очередь, позволило цветковым растениям, распространять свои семена не только с помощью ветра, а также с помощью животных и птиц, саморазбрасыванием и другими способами.
Семена покрытосеменных содержат большое количество запасных питательных веществ. Они содержатся либо в частях зародыша (обычно в семядолях), куда они перетекают в процессе созревания из эндосперма, либо остаются в эндосперме. Эндосперм — это особая ткань цветковых растений, которая представляет собой результат двойного оплодотворения. После опыления один спермий оплодотворяет яйцеклетку (образуется зигота), а второй так называемую центральную клетку семязачатка. В результате центральная клетка становится триплоидной (3n, содержит тройной набор хромосом). Ее деление формирует эндосперм, в которой притекает запас питательных веществ для зародыша растения, развивающегося из зиготы.
Эти и другие особенности в совокупности дали возможность покрытосеменным заселить новые места обитания, лучше приспособиться к ним. Цветковые оказались более прогрессивной группой растений, чем папоротники и голосеменные.
Читайте также: