В чем отличие между столбчатой и губчатой паренхимой листа чем обусловлено их расположение
Добавил пользователь Евгений Кузнецов Обновлено: 19.09.2024
Клетки имеют вытянутую форму, плотно сомкнуты, без межклетников, расположены в два слоя. Это столбчатая паренхима. Здесь в основном происходит фотосинтез.
Более округлые клетки с крупными межклетниками - губчатая паренхима. У нее в некоторых клетках можно заметить отложения оксалата кальция, а также крупные разветвленные механические клетки - склереиды, выполняющие опорную функцию.
Листья, у которых мезофилл дифференцирован на столбчатую и губчатую паренхиму, называют дорсовентральными.
вoва сам пиши, тебе википeдия зачем
Ри определенных условиях пища может быть причиной различных заболеваний микробной и немикробной природы. Нарушение санитарно-противоэпидемических и гигиенических норм и правил может привести к распространению пищевым путем некоторых инфекционных заболеваний (зоонозных и острых кишечных инфекций), к вспышкам пищевых отравлений микробной природы, к глистным заболеваниям.
Пищевые отравления могут возникать в результате попадания в пищу вредных примесей различного происхождения. Особенно актуально предупреждение загрязнения пищевых продуктов тяжелыми металлами, средствами химической защиты растений и другими химическими компонентами, попадающими во внешнюю среду в результате деятельности человека.
Вредные свойства пища может приобретать и в процессе приготовления в результате неправильного использования пищевых добавок, посуды, оборудования или при нарушении правил тепловой обработки продуктов за счет образования токсических и канцерогенных веществ.
Профилактика инфекционных и глистных заболеваний, пищевых отравлений в соответствии с законодательством должна осуществляться на всех предприятиях, производящих и реализующих пищевые продукты и пищу.
К острым кишечным инфекциям относится брюшной тиф, паратифы А и В, дизентерия, холера, сальмонеллез, инфекционный гепатит, иерсиниоз и др.

Справочник по биологии.
Основной тканью в сформировавшемся листе является мезофилл (рис. 13), обычно выполняющий фотосинтетическую функцию. Он представляет собой паренхимную ткань, состоящую из клеток, содержащих хлорофилл, богатую межклетниками. У большинства видов покрытосеменных растений-мезофилл дифференцирован на палисадную и губчатую паренхиму.

Палисадная паренхима состоит из клеток, удлиненных в направлении, перпендикулярном поверхности листа, и имеющих вид столбиков. Иногда эту ткань называют столбчатой паренхимой. Губчатая паренхима получила свое название в связи с тем, что она пронизана Системой хорошо выраженных межклетников.
Клетки палисадной паренхимы имеют призматическую форму (у некоторых растений неправильную), длина их в большинстве случаев превосходит ширину в несколько раз. Для губчатой паренхимы характерно наличие клеток самой разной формы (изодиаметрических, удлиненных перпендикулярно или параллельно поверхности), Палисадная паренхима у большинства растений располагается с верхней стороны листа, однако у ксерофитов — часто по обеим сторонам листа, а губчатая сильно редуцирована или отсутствует.
По характеру расположения палисадной паренхимы различают лист дорсивентральный — палисадная паренхима расположена на одной стороне листа — и изолатеральный — на обеих сторонах листа. Различают также листья центрического типа, когда мезофилл верхней и нижней сторон одинаков. Такое строение встречается у цилиндрических листьев и представляет собой модификацию изолатерального листа.
Считают, что наиболее высоко специализированным типом ткани, приспособленной к выполнению функции фотосинтеза, является палисадная паренхима. Это подтверждается большим количеством хлоропластов в дифференцированных клетках палисадной паренхимы по сравнению с губчатой.
Ультраструктура пластид в палисадных клетках отличается сильным развитием и уплотнением тилакоидов, в губчатой ткани хлоропласты имеют более рыхлую структуру. Это объясняется тем, что в онтогенезе дифференцировка губчатой ткани листа заканчивается рано, а пластидный комплекс в палисадной ткани формируется пока растет лист, и синхронно с этим нарастает активность фотосинтеза. Для палисадной ткани характерен также более высокий уровень синтеза белка, что соответствует росту клеток и новообразованию пластид на протяжении всего периода роста в отличие от губчатой ткани, где эти процессы прекращаются рано. Высокая степень специализации палисадной ткани, приспособленной к выполнению функции фотосинтеза, проявляется также в ее связи с системой межклетников, в ультраструктуре пластид.
Существует и другая точка зрения: губчатая ткань является основной ассимилирующей тканью листа, Палисадная паренхима лишь защищает губчатую от избыточного освещения. Низкая фотосинтетическая активность палисадной паренхимы связана с фотоинактивацией пластид в палисадных клетках и ограничением диффузии СО2 к ним.
Считают, что губчатая паренхима в большей степени, чем палисадная, специализирована к транспорту ассимилятов из листа, потому что в ней относительно интенсивнее синтезируется сахароза; в палисадной паренхиме в основном синтезируется крахмал. Этому положению соответствует и тот факт, что хорошо развитая наружная флоэма сосудисто-волокнистых пучков обращена вниз, к губчатой паренхиме. Очевидно, эти ткани включаются в фотосинтез листа параллельно как автономные и выполняющие специализированные функции.
Изучение структуры ассимиляционной ткани в зависимости от расположения листьев на стебле, а также от возраста показало, что у расположенных выше листьев размеры межклетников значительно меньше как в палисадной, так и в губчатой паренхиме. Это свойство особенно четко выражено в верхушках верхних листьев, где губчатая паренхима приобретает вид компактной ткани, в которой практически нет межклетников. Изменяется также толщина губчатой паренхимы: она значительна в листьях нижних ярусов, уменьшается в средних и снова увеличивается на верхушке. Толщина листовой пластинки и палисадной паренхимы почти в 3—5 раз меньше у одногодичных приростов. Эта особенность в строении разновозрастных листьев, естественно, отражается и на их функции.
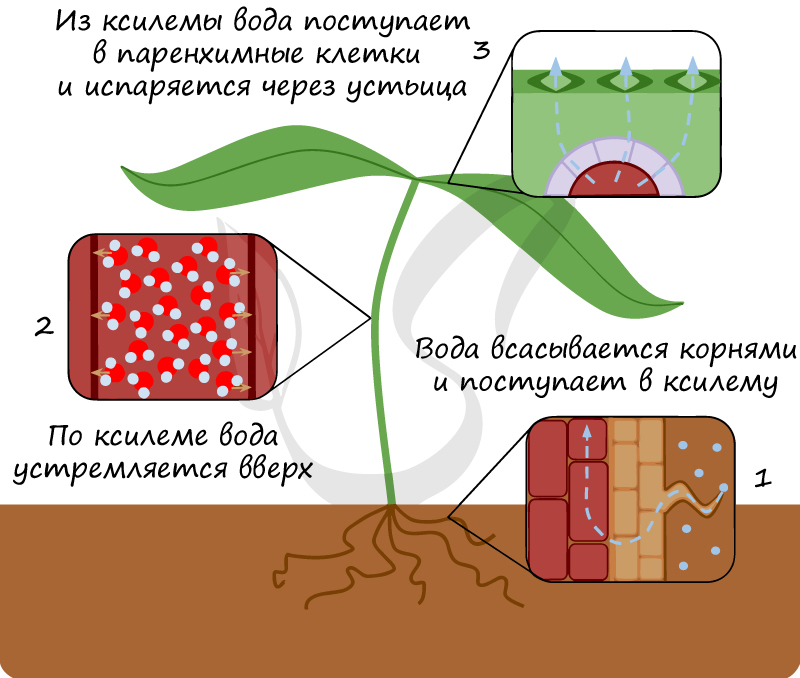
Транспортную функцию в листе выполняет проводящая система — жилки. Жилки — полифункциональные образования: они снабжают лист водой, минеральными и органическими веществами, притекающими из корня; обеспечивают отток ассимилятов, разгружая лист от продуктов фотосинтеза; выполняют механическую функцию, создавая опорный скелет листа и укрепляя его мякоть. Длина сети жилок зависит от многих внешних и внутренних факторов.
Передвижение веществ в листе происходит по флоэме и ксилеме. В наиболее крупных жилках листьев они образуют один или несколько пучков, располагающихся в виде кольца, полукольца или беспорядочно. Основным проводящим элементом ксилемы всех жилок за исключением конечных являются сосуды (трахеи), в самых маленьких конечных жилках — трахеидах — одна, две или группа. Трахеи и трахеиды в зрелом состоянии представляют собой более или менее удлиненные клетки с одревесневшими оболочками, лишенные протопластов. Трахеиды — неперфорированные клетки, имеющие пары пор на общих стенках. Трахеи — полые трубки, состоящие из ряда члеников, соединенных между собой. Перфорации у члеников сосудов обычно располагаются на их поперечных стенках, иногда на боковых. Вода, двигаясь по сосуду, свободно проходит через перфорации, тогда как в трахеидах она должна проходить через тонкие поровые мембраны. При росте ксилемы средняя площадь образующихся сосудов значительно увеличивается на ранних этапах. Число сосудов у некоторых растений (фасоли) продолжает возрастать и после прекращения роста листовой пластинки. Развитие ксилемы зависит главным образом от потребности листа в воде.
Основными проводящими элементами флоэмы являются ситовидные трубки и сопутствующие им клетки-спутники Ситовидная трубка состоит из клеток, характеризующихся своеобразным строением протопластов и наличием так называемых ситовидных полей — участков клеточной стенки, пронизанных многочисленными отверстиями, через которые с помощью тяжей сообщаются протопласты соседних ситовидных элементов. Части клеточной стенки, несущие ситовидные поля, называются ситовидными пластинками. Строение ситовидных Трубок (отсутствие ядра, наличие густой разветвленной сети фибрилл, отсутствие тонопласта, в результате чего протоплазма сильно оводнена) указывает на их большую специализацию, связанную, очевидно, со способностью к активному транспорту веществ.
Клетки-спутники — паренхимные клетки, образующиеся из той же меристематической клетки, что и связанная с ними клетка ситовидной трубки. У многих растений ситовидные элементы все время сопутствуют трахеидам, а иногда даже оказываются расположенными дальше их. В конечных ответвлениях жилок флоэмная часть может содержать только паренхиму.
Основным признаком, характеризующим проводящую систему листа, является ее связь с мезофиллом. Между мезофиллом и проводящей системой находится так называемое свободное пространство, или апопласт. Очевидно, с помощью сети проводящих пучков возможны адсорбция Сахаров и других ассимилятов из свободного пространства и концентрация их во флоэме для отвода из листа. Продолжением проводящей системы листьев является проводящая система черешков.
В листе, как и во всем растении, различают дальний транспорт веществ (происходит по флоэме и ксилеме), обеспечивающий координацию функций всего растения, и внутриклеточный, обеспечивающий передвижение веществ в клетке и осуществляющийся в основном с помощью цитоплазматических мембран. Обе системы транспорта функционируют в растении одновременно и тесно связаны между собой. В наиболее типичных случаях транспорт метаболитов начинается в фотосинтезирующих хлоропластах, из хлоропластов распространяется в цитоплазму, затем проходит через ряд паренхимных клеток и, преодолев концентрационный порог, продолжается в системе клеток флоэмы, по которым ассимиляты передвигаются к потребляющим тканям. Здесь метаболиты снова переходят в сферу паренхимного транспорта и заканчивают свое направленное перемещение сосредоточением в запасающих или потребляющих клетках.
Кроме того, в растении функционирует ксилемный транспорт, с помощью которого в основном происходит распределение воды и элементов корневого питания. Обе транспортные системы, вытянутые параллельно и прижатые Друг к другу, могут частично взаимодействовать путем обмена подвижными веществами.
Опорную функцию листа могут выполнять сами сосуды, так как они представляют собой мертвые образования с лигнифицированными стенками, а также механические волокна пучков.
Лист — орган специфический, и опорные структуры его не так хорошо развиты, как в стебле. Интенсивно развитая проводящая система является хорошей опорой для мезофилла. Дополнительную опорную функцию в листе несут колленхима, расположенная под эпидермисом над крупными жилками или по краю пластинки, склереиды, встречающиеся иногда в толще мезофилла, обособленные тяжи склеренхимы, имеющиеся у некоторых растений (пальм).
Опорную функцию может выполнять и покровная ткань листа — эпидермис (см. рис. 13), Плотно сомкнутые клетки, часто пропитанные кутином, иногда в различной степени лигнифицированные или инкрустированные кремнеземом (например, у злаков) и имеющие на наружной поверхности плотную кутикулу, представляют наружный скелет листа. Однако эта функция эпидермиса листа не главная. Плотно примыкая друг к другу, эпидермальные клетки создают покров, предохраняющий растение от иссушения, механических повреждений, проникновения внутрь насекомых, грибов-паразитов, бактерий, вирусов. Чем толще слой эпидермиса, тем сильнее он поглощает ультрафиолетовые лучи, защищая растение иногда даже за счет разрушения клеток самого эпидермиса.
Основная функция покровной ткани — защитная. Вместе с тем эпидермис легко проницаем для газов (СО2 и О2), воды, т. е. способен не только защищать лист, но и регулировать его газообмен, участвовать в фотосинтезе, поглощать воду и минеральные вещества. Полифункциональность, естественно, сказалась на строении ткани, включающей специализированные образования.
По характеру выполняемых функций, строению и происхождению эпидермис состоит из трех групп клеток: основные эпидермальные клетки, трихомы и устьица.
Основные эпидермальные клетки обычно плотно сомкнуты друг с другом, межклетники встречаются только в эпидермисе лепестков. Форма правильная, удлиненная, часто клетки имеют волнистые стенки. Оболочки клеток неравномерно утолщены, наиболее утолщена наружная стенка. Клеточные оболочки пропитаны кутином. Кутин может отлагаться на наружной поверхности клеток в виде самостоятельного слоя — кутикулы.
Трихомы представлены в листе защитными кроющими и железистыми волосками. По структуре их можно разделить на одноклеточные (простые или ветвистые) и многоклеточные (один или несколько рядов клеток). Закладываются они в результате разрастания одной из эпидермальных клеток, которая образует как бы выступ над поверхностью эпидермиса. Этот выступ удлиняется, а в случае развития многоклеточного волоска делится в различных направлениях. Волоски недолговечны, нередко они отмирают с окончанием роста листа или даже раньше, однако иногда функционируют дольше.
В зависимости от строения трихом, их размеров, числа на единицу площади, ориентации выделяют определенные типы опушения листа: войлочное — густое из ветвистых волосков (лист коровяка); мохнатое— извилистые волоски, не имеющие определенной ориентации (лист бегонии королевской); шелковистое — тонкие прямые волоски, обычно ориентированные в одном направлении; железистое — железистые трихомы в отличие от простых выделяют различные секреты (эфирные масла, слизи, смолы, растворы солей).
Устьица — система высокоспециализированных клеток, резко отличающихся по строению от основных клеток эпидермиса. Устьице состоит из двух замыкающих клеток, между которыми находится устьичная щель. У большинства двудольных растений замыкающие клетки имеют бобовидную форму, у однодольных более или менее удлиненную. Замыкающие клетки могут располагаться на одном уровне с соседними эпидермальными клетками, выступать над ними или быть погруженными, располагаясь ниже поверхности эпидермиса. Замыкающие клетки часто окружены двумя или большим числом клеток, отличающихся по форме от основных клеток эпидермиса. Эти клетки называются около-устьичными, или побочными. Совокупность околоустьичных и замыкающих клеток составляет устьичный аппарат, или устьичный комплекс.
Устьица расположены обычно на нижней стороне листа; у многих ксерофитов — и на нижней и на верхней. Исключение составляют злаки, у которых устьица находятся на верхней стороне листа. Подобное строение имеют листья плавающих водных растений. В участках эпидермиса, расположенных над проводящими пучками, устьиц обычно нет, они находятся лишь над клетками ассимиляционной ткани.
Среднее число устьиц на 1 мм 2 поверхности листа — примерно 100—300. Число устьиц непостоянно не только для различных ярусов листьев в пределах одного и того же растения, но может быть еще более непостоянно и в пределах одного и того же листа, изменяясь как от основания к верхушке, так и от края листа к средней его жилке, а также в процессе онтогенеза. Однако бывают исключения, когда число устьиц довольно равномерно распределено по всей пластинке листа.
Характерной особенностью строения замыкающих клеток устьиц является неравномерное утолщение их оболочек. Эта особенность связана с механизмом устьичных движений, который связан также с изменением их тургорного давления. Предполагают, что изменение тургорного давления устьиц происходит с помощью определенных сократительных структур и сопровождается значительной затратой энергии; в регуляции осмотического давления участвуют не сахара, как считали ранее, а органические кислоты, сахара являются лишь первичными промежуточными продуктами для образования осмотически активных веществ; в регуляции осмотического давления устьиц значительную роль играют одновалентные катионы, которые при открывании устьиц поступают из соседних клеток в замыкающие клетки против градиента концентрации. Некоторые исследователи рассматривают калий как вещество, играющее основную роль в регуляции осмотического потенциала устьичных клеток. Механизм открывания устьичной щели связан с затратой энергии. Эта энергия, очевидно, используется для активного транспорта катионов в вакуоли замыкающих клеток.
В итоге механизм движения устьиц можно представить следующим образом: при открывании устьичной щели одновалентные катионы (главным образом калий) против градиента концентрации поступают в вакуоль замыкающих клеток. Осмотический потенциал клеток возрастает, в них поступает вода, устьице открывается. Когда же механизм, поддерживающий высокую концентрацию солей в вакуолях замыкающих клеток, отключается, осмотически активные вещества выходят из вакуолей в цитоплазму, накапливаются в ней или выводятся из клетки. Осмотическое давление в клетке падает, снижается тургор, устьица закрываются.
Электронейтральность замыкающих клеток при открытых устьицах поддерживается в основном за счет образования органических анионов, которые в замыкающих клетках представлены главным образом анионами яблочной кислоты. Очевидно, именно эти анионы обусловливают закрывание устьиц, так как, если они не выводятся быстро из цитоплазмы в вакуоль или не обмениваются на ионы калия, цитоплазма приобретает кислую реакцию. Абсцизовая кислота способна блокировать активное вымывание ионов водорода из замыкающих клеток, тем самым препятствуя проникновению ионов калия в них, что в итоге приводит к закрыванию устьиц. Несмотря на то что в расшифровке механизма регуляции тургорного давления достигнуты определенные успехи, эта сложная проблема еще не решена.
Богданова, Т.Л. Справочник по биологии/ Т.Л. Богданова [и д.р.]. – К.: Наукова думка, 1985.- 585 с.

.
.
.
.
.
Показать полностью.
.
.
.
.
.
.
.
столбчатый мезофилл - фотосинтезирующая паренхима, располагающаяся на верхней стороне листа.клетки расположены плотно друг к другу, вытянутой формы. основная ф. - фотосинтез.
губчатый мезофилл - фотосинтезирующая паренхима, располог. на нижней стороне листа. крупные межклетники. клетки округлой формы. основная функция - транспирация, газообмен, транспорт веществ, фотосинтез - в меньшей степени, ибо мало хлоропластов.
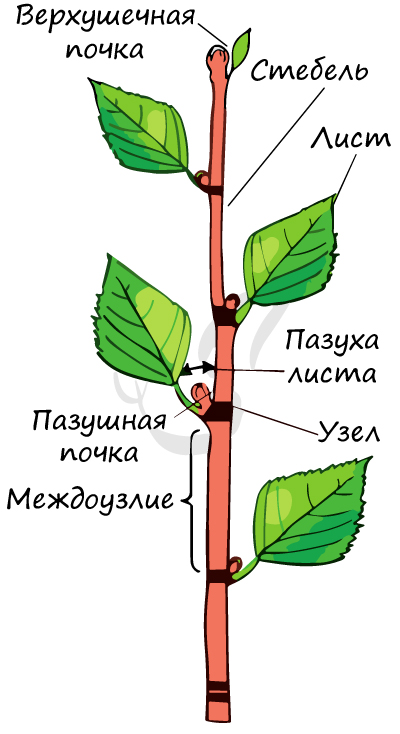
Лист - вегетативный орган растения, располагающийся на побеге. Место расположения листа на побеге называется узел. Узел (лат. nodus) — участок побега (стебля) растения, от которого отходят боковые органы (ветви, листья, почки, придаточные корни и другие.)
Строение и функции листа
Основная ткань пластинки листа - мезофилл. Выделяют столбчатый и губчатый мезофилл, функции которых различны.
Благодаря наличию хлоропластов в клетках столбчатой ткани мякоти листа происходит процесс фотосинтеза, в результате которого образуется большое количество органических веществ, доставляемых флоэмой в разные части растения. Вообразите следующую информацию в виде 3D-модели: проводящая система листа является продолжением проводящей системы стебля, в месте узла происходит отхождение сосудисто-волокнистого пучка в направлении листа.
В губчатой ткани листа расположены межклетники, вход в которые открывают устьица. Здесь происходит газообмен между организмом растения и внешней средой, заключающийся в процессах дыхания и фотосинтеза. Крайне важно разделить два понятия: фотосинтез и дыхание.
Не удивляйтесь тому, что растения поглощают кислород в процессе дыхания. Все живые клетки аэробных организмов находятся в процессе дыхания постоянно, днем и ночью. Запомните, что дыхание это поглощение кислорода и выделение углекислого газа. В ходе светозависимой фазы фотосинтеза напротив, кислород выделяется как ненужный побочный продукт, а углекислый газ поглощается клетками.

Осуществляется через устьица в эпидермисе (кожице).
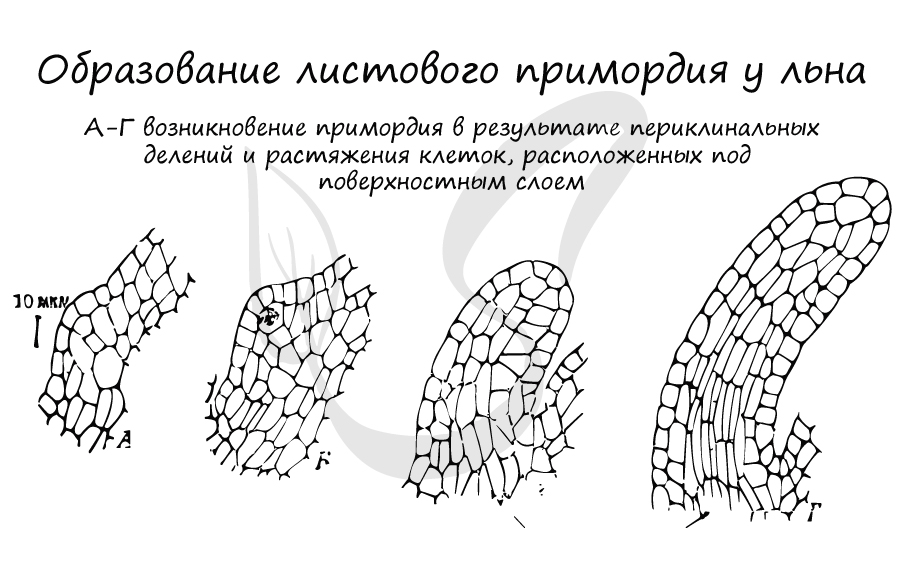
Самые первые листья растения - зародышевые листья (семядоли или семенодоли), которые развиваются у зародыша ещё в семени. В дальнейшем листья формируются из примордиев - нерасчленённых зачатков листьев в виде бугорков или валиков на конусе нарастания побега.
Основные части листа
Это расширенная нижняя часть листа. У некоторых растений оно, разрастаясь, преобразуется в незамкнутую или замкнутую трубку, которую называют листовое влагалище.
Выполняет главные функции листа - газообмен и фотосинтез, в основании пластинка сужается и переходит в стеблевидный черешок.
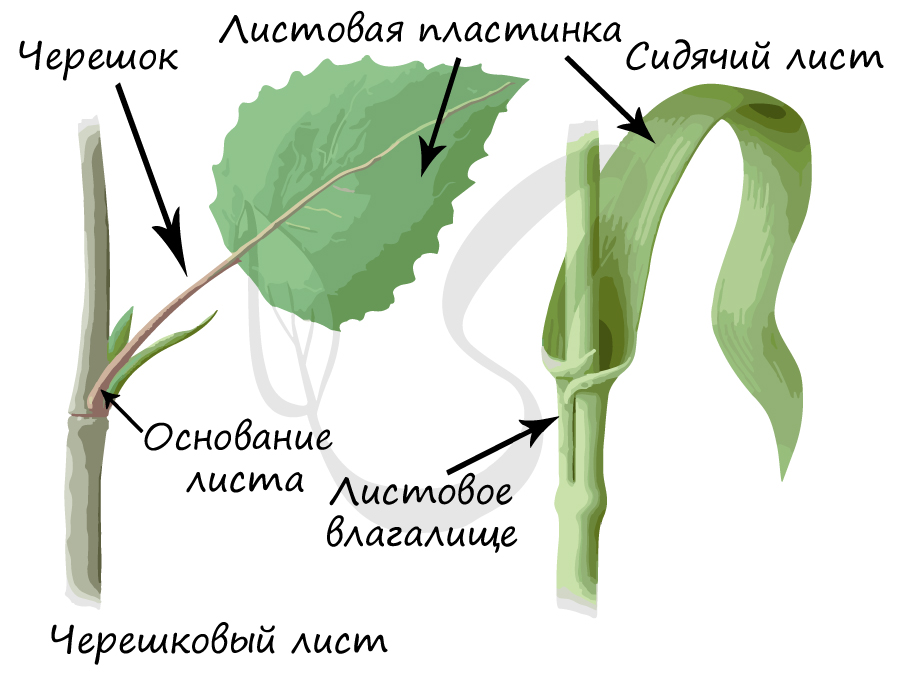
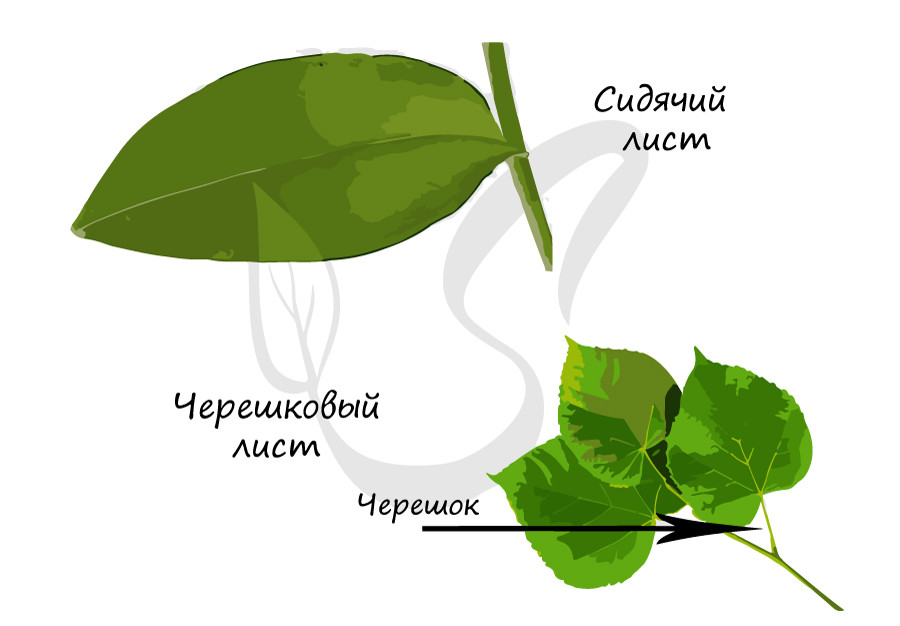
Это тонкая стеблевидная часть листа, идущая от листовой пластинки к узлу побега.
Меняя свою форму, черешок смещает листовую пластинку. Таким образом, основная функция черешка - обеспечить как можно большую освещенность листовой пластинки, вынести листья к свету. Именно так и создается листовая мозаика - расположение листьев на растении, при котором они не затеняют друг друга. Листья с черешками называются черешковыми, без черешка - сидячими.
Выросты листообразной формы, расположенные у основания листа. Они могут срастаться со стеблем или быть свободно расположенными. У многих растений прилистники отсутствуют в принципе, или образуются, но рано отмирают.
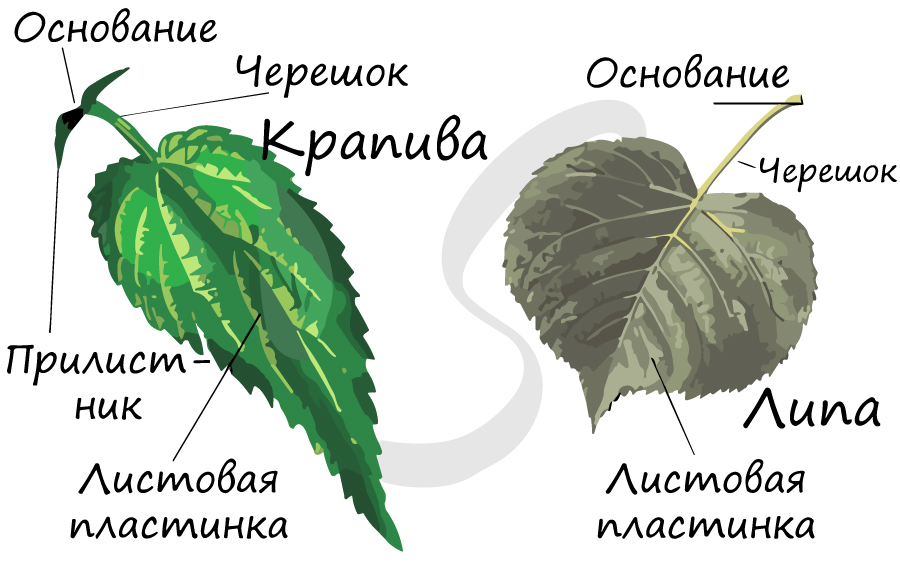
Лист называют полным, если в составе его элементов имеется пластинка, основание, прилистники и черешок. Полные листья характерны для многих широко известных растений: рябина, дуб, черемуха, роза.
Лист называют неполным, если у него отсутствует черешок (сидячий лист), прилистники или пластинка. Сидячие листы, лишенные прилистников, имеют лен, гвоздика, алоэ. Отсутствуют прилистники также на листьях картофеля, сирени, капусты. В редких случаях лист может не иметь листовой пластинки, тогда ее функции перенимают черешок - у акации, прилистники - у чины безлисточковой.

Жилкование листьев
-
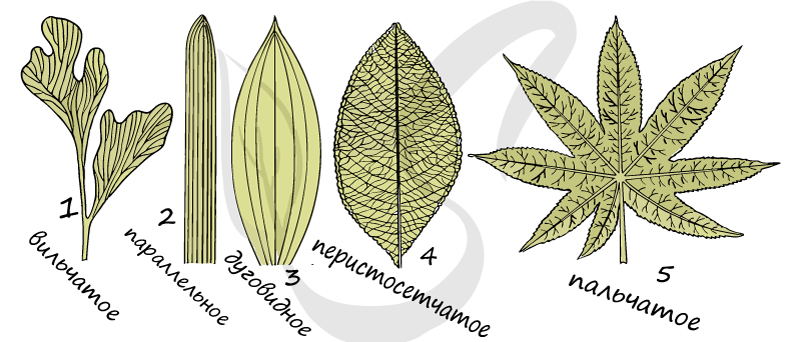
Вильчатое (дихотомическое) жилкование
Встречается у многих папоротниковидных растений и примитивных семенных, при вильчатом жилковании жилки делятся дихотомически (одна жилка разделяется на две жилки).
При таком типе жилкования крупные жилки проходят вдоль листовой пластинки параллельно друг другу. Характерно для злаковых растений.
Отличается наличием крупных жилок, которые подобно дуге изогнуты вдоль листовой пластинки. Характерно для однодольных.
Для этого типа характерна выраженная центральная (главная) жилка, от которой в стороны отходят более тонкие боковые ветви. Имеется у дуба черешчатого, черемухи обыкновенной.
Такой тип жилкования отличается наличием нескольких примерно одинаковых по размеру крупных жилок, расходящихся веером по пластинке, при этом сходящихся в одной точке у ее основания. Имеется у манжетки обыкновенной, клена платановидного.
Форма листа
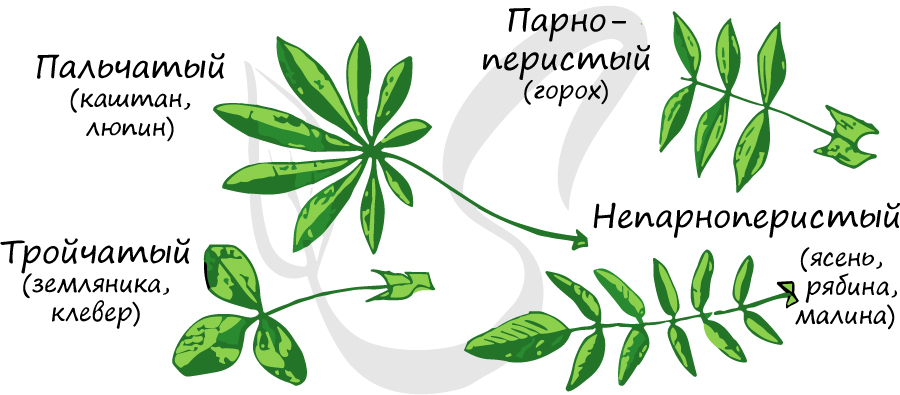
- Однолисточковые - у мандарина, лимона.
- Тройчатосложные - у земляники, клевера.
- Пальчатосложные, состоящие из множества листовых пластинок, у люпина, каштана конского.
Необходимо особо отметить, что есть сложные листья, у которых листочки расположены по всей длине рахиса - пункты 4 и 5.
- С цельной листовой пластинкой - сирень, береза, тополь, яблоня.
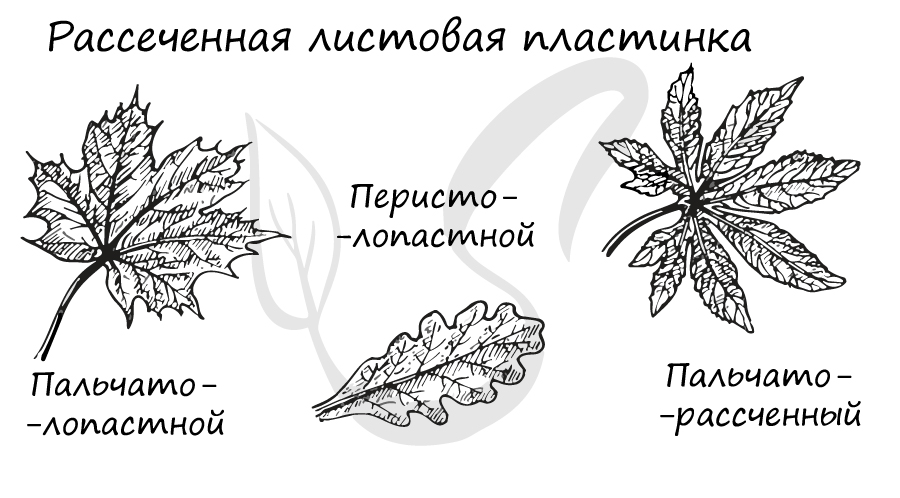
- С рассеченной (расчлененной) листовой пластинкой. Каждую отдельную часть простой пластинки называют сегментом. Здесь также имеется еще одно деление, по степени расчлененности листовых пластинок различают:
- Пальчтолопастную (перилопастную) - в случае если расчленение не превышает 1/3 всей поверхности листовой пластинки.
- Перистолопастную (перистораздельную) - расчленение не превышает половины (до 1/2) листовой пластинки.
- Пальчаторассеченную (перисторассеченную) - расчленение достигает главной жилки листа или основания листовой пластинки.
![Простой лист]()
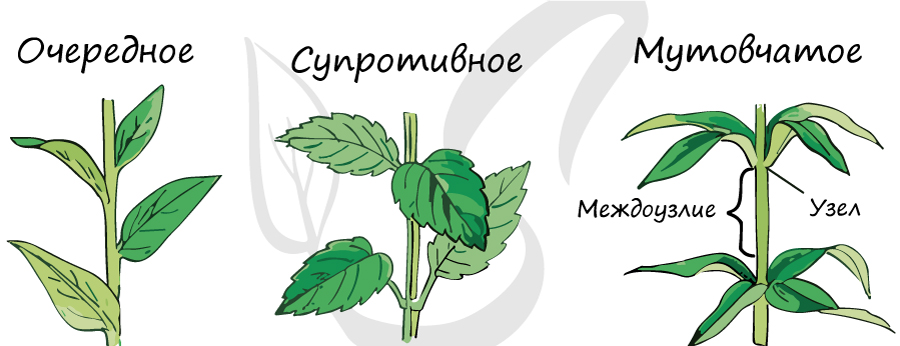
Листорасположение
- Очередное - от узла отходит только один лист. Имеется у березы, липы, дуба.
- Супротивное - на узле располагаются два листа, супротив (напротив) друг друга. Встречается у бузины, клена, калины.
- Мутовчатое - на узле стебля 3 и более листьев. Есть у вороньего глаза, ветренницы, элодеи.
![Листорасположение]()
Видоизменения листьев
Это интереснейшие приспособления, которые возникли в процессе приспособления растений к различным средам обитания. Они выполняют дополнительные функции, но главная их задача - это адаптация растения к условия среды.
Не все растения питаются автотрофно, для некоторых из них свойственен гетеротрофный тип питания. Известный пример росянка капская - насекомоядное растение. Ее лист покрыт липкой вязкой массой, которая выделяется волосками листа. Если насекомое садится на лист, то приклеивается к нему, волоски начинают сворачиваться, и насекомое оказывается в образовавшейся полости. После чего начинается выделение ферментов в полость и переваривание насекомого.
![Ловчие аппараты]()
Образования, которые выполняют опорную функцию. Цепляясь усиками за опору, растение занимает в пространстве вертикальное положение, растут вверх. Имеются у чины, гороха.
![Усики]()
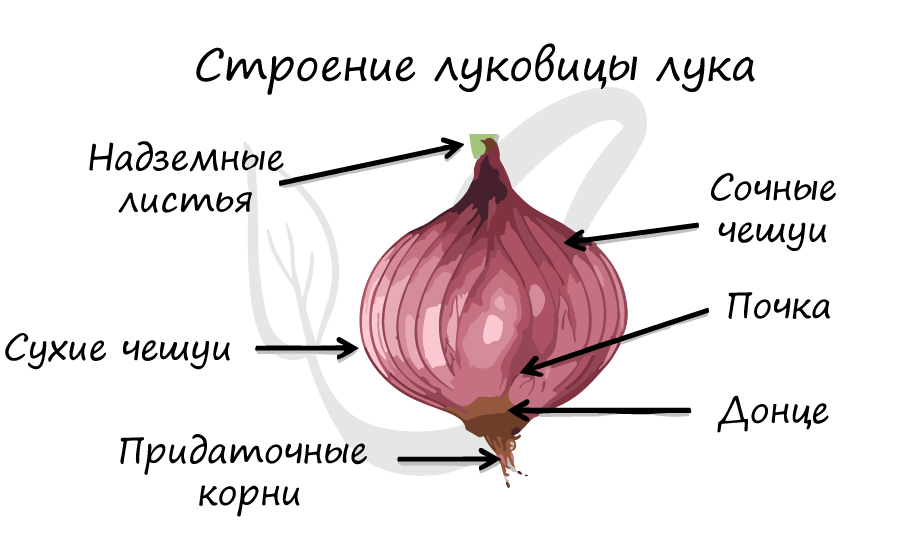
Выполняют различные функции. К примеру, чешуи почки защищают ее от механических повреждений, а листья у лука в луковице превращены в сочные чешуи, которые запасают питательные вещества.
![Чешуи]()
Ограждают растение от поедания его животными. Подобную защитную функцию выполняют колючки барбариса, кактуса.
![Колючки]()
Эти видоизменения листьев не утратили свою основную функцию, и приобрели дополнительную - запасание воды. Особенно актуальна эта функция для растений суккулентов, произрастающих в местах с засушливым климатом - алоэ, молодил, очиток.
![Сочные и толстые образования, запасающие воду]()
Хвоя - это видоизмененные листья голосеменных (хвойных) растений. Таким листьям, в отличие от неизмененных, нужно меньше питательных веществ и воды. Они способны противостоять холоду и засухе, при этом выполняя свою основную роль - обеспечение процесса фотосинтеза.
Вечнозелеными растениями является подавляющее большинство голосеменных, которые сохраняют хвою в течение 12 месяцев, не сбрасывая ее перед зимой. У отдельных видов, сосны долговечной, хвоя сохраняется до 45 лет.
![Хвоя]()
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: