
Сумма ежедневных показателей площади листьев посева за определенный период называется
Добавил пользователь Skiper Обновлено: 19.09.2024
В статье представлен процесс формирования фотосинтетического аппарата яровой пшеницы при применении биологических препаратов. Исследованиями установлено, что интенсивность нарастания листовой поверхности в течение всей вегетации достигала максимальных значений при инокуляции семян бактофосфином в смеси с ЖУСС-2 (жидкий удобрительно-стимулирующий состав) на фоне минеральных удобрений. При применении биопрепаратов интенсивность нарастания фотосинтетического потенциала посевов в течение вегетации яровой пшеницы возрастает за счет увеличения листовой поверхности. На характер формирования листовой поверхности яровой пшеницы минеральные удобрения и биопрепараты оказали непосредственное воздействие. Максимальные величины чистой продуктивности фотосинтеза наблюдались в фазе колошения, т.е. в период наибольшего развития листовой поверхности растений яровой пшеницы. Инокуляция семян способствует увеличению фотосинтетической деятельности растений в посевах и улучшению динамики ростовых процессов. Исследованиями установлено, что на характер формирования листовой поверхности яровой пшеницы минеральные удобрения и биопрепараты оказали непосредственное воздействие. Инокуляция семян биопрепаратами увеличивает данный показатель фотосинтетической деятельности, вероятно, за счет стимулирующего действия микроорганизмов, входящих в состав биопрепаратов, которая начинает проявляться с фазы кущения. Как при применении минеральных удобрений, так и без них, наибольшую площадь листьев имели растения на варианте бактофосфин + ЖУСС. Инокуляция семян способствует увеличению фотосинтетического потенциала посевов в течение вегетации за счет увеличения листовой поверхности. Инокуляция на фоне минеральных удобрений дает возможность формирования более высокого фотосинтетического потенциала посевов яровой пшеницы. При инокуляции семян яровой пшеницы биопрепаратами и препаратом ЖУСС-2 проявляется тенденция к повышению чистой продуктивности фотосинтеза как на неудобренном фоне, так и на фоне минеральных удобрений.

3. Мокроносов А.Т. Фотосинтетическая функция и целостность растительного организма [42-е Тимирязевские чтения]. – М.: Наука, 1983. – 63 с.
4. Мокроносов А.Т. Фотосинтез и продукционный процесс / А.Т. Мокроносов // Физиология растений на службе продовольственной программы СССР. – М.: Знание, 1988. – № 2. – С. 3–18.
6. Никитин С.Н. Влияние бактериальных удобрений на микрофлору почвы и урожайность яровой пшеницы // Агроэкологические аспекты повышения эффективности сельскохозяйственного производства: тез. докл. научно-практической конференции. – Пенза, 2001. – С. 127–129.
7. Ничипорович А.А. Некоторые принципы комплексной оптимизации фотосинтетической деятельности и продуктивности растений // Важнейшие проблемы фотосинтеза в растениеводстве. – М.: Колос, 1970. – С. 120–127.
8. Ничипорович А.А. Реализация регуляторной функции света и жизнедеятельности растений как целого и в его продуктивности. / А.А. Ничипорович. Фоторегуляция метаболизма и морфогенеза растений. – М., 1975. – 275 с.
9. Ничипорович А.А. Физиология фотосинтеза и продуктивность растений / А.А. Ничипорович. Физиология фотосинтеза. – М., 1982. – С. 7–34.
11. Ничипорович А.А. Фотосинтетическая деятельность растений в посевах / А.А. Ничипорович, Л.Е. Строганова, С.Н. Чмора, М.Н. Власова. – М.: Изд-во АН СССР, 1961. – 137 с.
12. Оценка эффективности микробных препаратов в земледелии / Под общ. ред. А.А. Завалина. – М.: РАСХН, 2000. – 82 с.
13. Федин М.А. Методика государственного сортоиспытания сельскохозяйственных культур / М.А. Федин. – М.: Изд-во Министерство сельского хозяйства СССР, 1985. – 285 с.
14. Шевелуха В.С. Периодичность роста сельскохозяйственных культур и пути ее регулирования. – М.: Колос, 1980. – 455 с.
Взаимосвязь растений в агроценозе носит непостоянный характер, зависящий от многих факторов. Главной задачей для получения высоких урожаев является создание такого посева, в котором бы максимально раскрывались потенциальные возможности фотосинтетической деятельности растений в агроценозе. Этого можно добиться при создании благоприятных условий для роста и развития растений.
Ряд авторов [3, 14] рассматривает рост растений как процесс дифференцирования организма за счет образования новых и увеличения старых элементов его структуры (молекул, клеток, тканей и органов), оказывающий решающее влияние на распределение, перераспределение и использование образовавшихся при фотосинтезе и метаболизме органических веществ, а также поглощение минеральных солей и воды, идущих на образование поверхности органов и тканей, их регенерацию и на запасные отложения. Рост растения суммирует в себе и венчает многие другие процессы жизнедеятельности растения, выражая в известной мере баланс процессов синтеза и распада веществ в организме при его взаимодействии с условиями внешней среды.
Рост растения и его биологическая продуктивность – результат, прежде всего, фотосинтетической деятельности, в ходе которой образуется до 95 % органических соединений. Поэтому рост растения, формативный, органообразовательный и рост как увеличение сухой биомассы начинается, главным образом, вслед за формированием фотосинтетической системы листа и осуществления процесса фотосинтеза. Лист как орган фотосинтеза является центром образования первичных продуктов, их метаболизации и эвакуации в органы запаса [2, 15].
Целью данной работы являлось выявление особенностей формирования фотосинтетической деятельности растений в посевах и динамика ростовых процессов при применении биологических препаратов и минеральных удобрений.
Материалы и методы исследования
Проводили следующие учеты и анализы:
– фенологические наблюдения согласно методике государственного сортоиспытания [13];
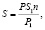
где S – общая площадь листьев одного растения, см2; S1 – площадь одной высечки, см2; P – общая масса листьев одного растения, г; P1 – масса высечек, г; n – число высечек. Зная густоту стояния растений на 1 га рассчитывалась площадь листьев на 1 га;
– содержание сухого вещества определялось отбором средней пробы измельченных растений. Из каждого образца отбирались навески по 50 г в двукратной повторности, фиксировались и высушивались в сушильном шкафу при температуре 105 °С до постоянной массы. Содержание сухого вещества определялось по формуле
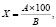
где А – масса навески после высушивания, г; В – масса навески до высушивания, г;
– чистая продуктивность фотосинтеза вычислялась по формуле Кидда, Веста и Бриггса:
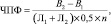
где ЧПФ – чистая продуктивность фотосинтеза; В1 и В2 – сухая биомасса пробы урожая в начале и конце учетного периода, г; В2 – В1 – прирост сухой массы за учетный период n дней, г; (Л1 + Л2)/0,5 – средняя работавшая площадь листьев за этот промежуток времени, м2; n – число дней.
Минеральные удобрения аммиачная селитра, двойной гранулированный суперфосфат и хлористый калий вносили под предпосевную культивацию. Обработка семян бактериальными препаратами и препаратом ЖУСС-2 (жидкий удобрительно-стимулирующий состав) проводили за 1–2 дня до посева. Повторность опыта 4-кратная. Учетная площадь делянок 42 м2 (2,1*20).
Результаты исследования и их обсуждение
Формирование фотосинтетического аппарата представляет сложный процесс. В ранние фазы роста и развития преобладают процессы новообразования и роста листьев, а в более поздние – процессы отмирания, связанные с усиленной транспортировкой пластических веществ в репродуктивные органы [9]. Накопление и запасание энергии в процессе фотосинтеза сопровождается накоплением биомассы, служащей структурным и энергетическим материалом, обеспечивающим существование растений [15].
Максимальные урожаи могут быть сформированы посевами с оптимальной площадью листьев, причем важно, чтобы она быстро нарастала до максимальной величины и долго удерживалась на достигнутом уровне без резкого снижения к концу вегетации, максимально поглощая солнечную радиацию [10]. У культур, потенциально способных к формированию очень большой листовой поверхности, к которым относятся, в частности, злаки [8], степень ее фактического развития, а тем самым и факторы фотосинтетической деятельности, лимитирующие урожайность, решающим образом зависят от условий выращивания. Если ресурсы влаги и питания недостаточны, то основным фактором, лимитирующим урожайность растений, является недостаточное развитие листовой поверхности, а на бедных почвах – и низкая производительность ее работы.
Основными показателями фотосинтетической деятельности растений являются: площадь листьев, фотосинтетический потенциал, чистая продуктивность фотосинтеза и коэффициент использования фотосинтетически активной радиации. Величина площади листьев слагается из площади листьев отдельных растений и в различной степени зависит от периода вегетации, погодных условий выращивания, сорта, густоты растений, обеспеченности их питательными веществами и т.д.
Исследования [4, 5] указывают, что более продуктивным может быть то растение, которое отличается менее интенсивным фотосинтезом, но больший процент ассимилятов использует на образование листьев и формирует большую ассимиляционную поверхность. Поэтому очень важно агротехническими мероприятиями в наиболее короткие сроки добиваться оптимальной площади листьев растений.
Имеется немало указаний в литературе, что с увеличением ассимиляционной площади листьев увеличивается урожай. Однако положительная связь этих двух процессов имеет предел, при котором большая листовая поверхность из-за взаимного затенения снижает интенсивность фотосинтеза, в результате чего увеличивается непродуктивная часть урожая и уменьшается продуктивная.
Наши исследования показывают, что в начале развития растений яровой пшеницы от появления всходов до фазы кущения только начинают проявляться различия вариантов по площади листьев. Площадь листьев в фазу кущения изменяется в пределах 7,5–8,9 тыс. м2/га, а на фоне минеральных удобрений 9,1–11,4 тыс. м2/га (табл. 1). При инокуляции семян наблюдается тенденция к увеличению ассимиляционной поверхности листьев, особенно это заметно при применении бактофосфин + ЖУСС-2. По-видимому, в данный период действие биопрепаратов только начинает проявляться [6].
Влияние биопрепаратов на ассимиляционную поверхность листьев, тыс. м2/га (среднее за 3 года)
Урожай создается в процессе фотосинтеза, когда в зеленых растениях образуется органическое вещество из диоксида углерода, воды и минеральных веществ. Энергия солнечного луча переходит в энергию растительной биомассы. Эффективность этого процесса и, в конечном счете, урожай зависят от функционирования посева как фотосинтезирующей системы.
В полевых условиях посев (ценоз) как совокупность растений на единице площади представляет собой сложную динамическую саморегулирующуюся фотосинтезирующую систему. Эта система включает в себя много компонентов, которые можно рассматривать как подсистемы; она динамическая, так как постоянно меняет свои параметры во времени; саморегулирующаяся, так как, несмотря на разнообразные воздействия, посев изменяет свои параметры определенным образом, поддерживая гомеостаз.
Такую систему характеризуют новые свойства по сравнению с отдельным растением. Так, для отдельного растения увеличение площади питания и связанное с этим улучшение освещенности приводят к повышению его семенной продуктивности, а для ценоза важна оптимальная густота растений. Условия для максимальной продуктивности отдельного растения и ценоза как системы не совпадают.
Исследованию фотосинтеза ценозов была посвящена Международная биологическая программа. Изучению фотосинтетической деятельности растений в посевах тесно связано с теорией получения высоких урожаев и возможностью управления формированием урожая. Методические основы ее изучения разрабатывали многие ученые. Большой вклад внес А. А. Ничипорович.
Управление формированием урожая весьма сложно, так как растения в ценозе, изменяясь в процессе вегетации, взаимодействуют с другими сложными системами – микроорганизмами почвы, возбудителями болезней, сорняками, вредителями. Многие факторы среды – температурный режим, осадки и другие – практически невозможно контролировать. Однако на основании анализа природно-климатических факторов можно подобрать сорта, адаптированные к конкретным условиям, разработать технологию их возделывания. Многие факторы поддаются регулированию. Можно изменить условия минерального питания, воздействовать на сорняки, вредителей болезни. Управление процессом формирования урожая ведут на основе систематического контроля за развитием растений и направления хода фотосинтетической деятельности посевов в соответствии с заранее заданными параметрами.
Фотосинтетически активная радиация (ФАР)
Необходимое условие фотосинтеза – энергия солнечной радиации. Следует создавать такие посевы, в которых листья поглощали бы энергию солнца с возможно более высоким коэффициентом полезного действия для создания наибольшей биомассы и сосредоточения ее в хозяйственно ценной части урожая – семенах, клубнях, корнеплодах и т. п.
В процессе фотосинтеза принимает участие не вся солнечная энергия, а только ее видимая часть – фотосинтетически активная радиация (ФАР) с длиной волн – 380-720нм (нанометр). Эти лучи поглощаются хлорофиллом и являются энергетической основой фотосинтеза. Энергия ФАР составляет около 50% общей энергии солнечной радиации. Инфракрасная часть солнечного спектра, составляющая также около 50% общей энергии солнца, не участвует в фотохимических реакциях фотосинтеза. Эти лучи поглощаются почвой, от которой нагреваются приземный слой воздуха и сами растения, при этом усиливаются транспирация и испарение влаги с поверхности почвы. Количество ФАР, падающее на единицу поверхности почвы в среднем по месяцам года и по декадам месяца, определено для различных географических зон и приведено в соответствующих справочниках.
Объективным показателем величины урожая (высокий, средний, низкий) может служить коэффициент использования ФАР. Хорошие урожаи соответствуют 2-3% использования ФАР. При возделывании сортов интенсивного типа и оптимизация всех процессов формирования урожая возможна аккумуляция в урожае 3,5-5% ФАР и более.
Поступление солнечной энергии за вегетационный период зависит от географической широты. Так, если сравнивать северные и южные районы РФ, то приход ФАР за возможный вегетационный период различается более чем в 2 раза. Кроме того, можно отметить, что на плодородных почвах Краснодарского края при достаточном количестве тепла и влаги значительно легче сформировать посев, который в среднем за вегетацию усваивает 3% ФАР, чем в Тверской области на малоплодородных почвах и при недостатке тепла.
Показатели фотосинтетической деятельности посевов
Посев представляет собой оптическую систему, в которой листья поглощают ФАР. В начальный период развития растений ассимиляционная поверхность невелика и значительная часть ФАР проходит мимо листьев, не улавливается ими. С повышением площади листьев увеличивается и поглощение ими энергии солнца. Когда индекс листовой поверхности (величина, показывающая, во сколько раз площадь листьев превышает ту площадь, на которой находятся растения; так, если индекс листовой поверхности равен 4, то площадь листьев составляет – 40тыс м?/га, или 4м?/м?) составляет – 4-5, т. е площадь листьев в посеве – 40-50тыс м?/га, поглощение ФАР листьями посева достигает максимального значения – 75-80% видимой, 40% общей радиации. При дальнейшем увеличении площади листьев поглощение ФАР не повышается.
В посевах, где ход формирования площади листьев оптимальный, поглощение ФАР может составить в среднем за вегетацию – 50-60% падающей радиации. Поглощенная растительным покровом ФАР – энергетическая основа для фотосинтеза. Однако в урожае аккумулируется только часть этой энергии. Коэффициент использования ФАР обычно определяют по отношению к падающей на растительный покров ФАР. Если в урожае биомассы в средней полосе РФ аккумулировано – 2-3% прихода на посев ФАР, то сухая масса всех органов растений составит – 10-15т/га, а возможная урожайность – 4-6т зерна/га. В изреженных посевах коэффициент использования ФАР составляет всего – 0,5-1%.
При рассмотрении посева как фотосинтезирующей системы урожай сухой массы, создаваемый за вегетационный период, или его прирост за определенный период зависит от величины средней площади листьев, продолжительности периода и чистой продуктивности фотосинтеза за этот период.
где У – урожайность сухой биомассы, т/га; ФП – фотосинтетический потенциал, тыс. м?*дни/га; ЧПФ – чистая продуктивность фотосинтеза, г/(м?*дни).
Фотосинтетический потенциал рассчитывают по формуле:
где Sc – средняя за период площадь листьев, тыс. м?/га; Т – продолжительность периода, дни.
Основные показатели для ценоза, как и урожайность, определяют в расчете на единицу площади – 1м? или 1га. Так, площадь листьев измеряют в тыс. м?/га. Кроме того, пользуясь таким показателем, как индекс листовой поверхности.
Основную часть ассимиляционной поверхности составляют листья, именно в них осуществляется фотосинтез. Фотосинтез может происходить и в других зеленых частях растений – стеблях, остях, зеленых плодах и т.п., однако вклад этих органов в общий фотосинтез обычно небольшой. Принято сравнивать посевы между собой, а также различные состояния одного посева в динамике по площади листьев, отождествляя ее с понятием “ассимиляционная поверхность”.
Динамика площади листьев в посеве подчиняется определенной закономерности. После появления всходов площадь листьев медленно повышается, затем темпы нарастания увеличиваются. К моменту прекращения образования боковых побегов и роста растений в высоту площадь листьев достигает максимальной за вегетацию величины, затем начинает постепенно снижаться в связи с пожелтением и отмиранием нижних листьев. К концу вегетации в посевах многих культур (зерновые, зернобобовые) зеленые листья на растениях отсутствуют.
Площадь листьев различных с/х растений может сильно варьировать в течение вегетации в зависимости от условий водоснабжения, питания, агротехнических приемов. Максимальная площадь листьев в засушливых условиях достигает всего – 5-10тыс м?/га, а при избыточных увлажнении и азотном питании она может превышать – 70тыс м?/га. Считается, что при индексе листовой поверхности 4-5 посев, как оптическая фотосинтезирующая система, работает в оптимальном режиме, поглощая наибольшее количество ФАР. При меньшей площади листьев часть ФАР листья не улавливают. Если площадь листьев больше 50тыс м?/га, то верхние листья затеняют нижние, их доля в фотосинтезе резко снижается. Более того, верхние листья “кормят” нижние, что не выгодно для формирования плодов, семян, клубней и т.д.
Динамика площади листьев показывает, что на разных этапах вегетации посев как фотосинтезирующая система функционирует неодинаково. Первые 20-30 дней вегетации, когда средняя площадь листьев составляет – 3-7тыс м?/га, большая часть Фар не улавливается листьями, и поэтому коэффициент использования ФАР не может быть высоким. Далее площадь листьев начинает быстро нарастать, достигая минимума. Как правило, это происходит у мятликовых в фазе молочного состояния зерна, у зернобобовых – в фазе полного налива семян в среднем ярусе, у многолетних трав – в фазе цветения. Затем площадь листьев начинает быстро снижаться. В это время преобладают перераспределение и отток веществ из вегетативных органов в генеративные.
На продолжительность этих периодов и их соотношение влияют различные факторы, в том числе агротехнические. С их помощью можно регулировать процесс нарастания площади листьев и продолжительность периодов. В засушливых условиях густоту растений, а следовательно, и площадь листьев намеренно снижают, так как при большой площади листьев усиливается транспирация, растения сильнее страдают от недостатка влаги, урожайность уменьшается.
Слишком большое разрастание площади листьев при достаточном водоснабжении также приводит к нежелательным результатам. Биомасса в этом случае растет довольно высокими темпами за счет вегетативных органов, однако условия формирования плодов и семян ухудшаются. К подобным результатам может привести и чрезмерное загущение растений. Для кормовых культур, у которых листья представляют хозяйственно ценную часть урожая (например травы), площадь листьев может достигать – 60-80тыс м?/га.
Таким образом, получение высоких урожаев связано с оптимальным ходом (графиком) нарастания площади листьев. Такие графики могут быть определены для каждой культуры и сорта в конкретных условиях их выращивания.
Формирование урожая зависит не только от величины площади листьев, но и от времени ее функционирования. Фотосинтетический потенциал (ФП) объединяет эти показатели. ФП может быть определен за любой период времени, например за декадные, межфазные периоды или в целом за вегетационный период. ФП за какой-либо период представляет сумму величин площади листьев за каждые сутки периода. Например, если площадь листьев в начале периода составляет – 20тыс м?/га, а через 10 дней – 28тыс м?/га, то ФП этого 10-дневного периода составляет – (20+28):2*10=240тыс м? * дни/га. ФП хорошо развитых посевов зерновых культур с вегетационным периодом 100-110 дней составляет за вегетацию – 2-2,5млн м?*дни/га. В южных районах для культур с длительным вегетационным периодом и при хорошем водоснабжении ФП может составить до 4млн м?*дни/га.
Чистая продуктивность фотосинтеза (ЧПФ) характеризует интенсивность фотосинтеза посева и представляет собой количество сухой массы растений в граммах, которое синтезирует 1м? листовой поверхности за сутки. В среднем за вегетацию у таких культур, как пшеница, ячмень, ЧПФ составляет – 5-7г/(м?*дни). У кукурузы ЧПФ обычно выше. ЧПФ, так же как и ФП, определяют за какой-либо период или в среднем за вегетацию:
где В2 и В1 – сухая масса растений с единицы площади в конце и начале периода.
ЧПФ варьирует в течение вегетации. В первый месяц вегетации ЧПФ выше, чем в последующий, так как в начале вегетации растения не затеняют друг друга, все листья хорошо освещены. В дальнейшем с увеличением площади листьев ЧПФ начинает уменьшаться в связи с затенением нижних листьев.
В начале вегетации нарастание биомассы идет медленно, затем темпы приростов увеличиваются. В конце вегетации, когда площадь листьев небольшая, суточные приросты биомассы также невелики. В это время идет перераспределение накопленных ассимилятов из листьев, стеблей и корней в генеративные органы.
Прирост биомассы за любой промежуток времени, в том числе за вегетацию, равен произведению ФП и ЧПФ. Если в среднем за 100 дней вегетации ЧПФ равнялось – 6г/(м?*дни), а ФП – 2млн м?*дни/га, то количество сухой массы составит – 12т/га.
Посев как фотосинтезирующая система наиболее производительно функционирует в период, когда площадь листьев близка к оптимальной – 30-50тыс м?/га. Если ЧПФ в это время равна – 5-7г/(м?*дни), то при площади листьев – 40тыс м?/га суточный прирост сухой биомассы составит – 200-280кг/га. Если период с такой средней площадью листьев продолжается 30 дней, то за это время прирост сухой массы составит – 6-8т/га. При рост биомассы за период, когда посев функционирует в оптимальном режиме, составляет более 70% максимального за вегетацию, хотя продолжительность этого периода – всего 30% общей вегетации культуры. Коэффициент использования ФАР в это время в 2-3 раза больше, чем в первый месяц после появления всходов, а также в конце вегетации.
При созревании в корнях и стеблях сосредотачивается 50-60% сухой массы растений, в основном представленной клетчаткой. В нашем примере из общей биомассы 12т/га на урожай зерна приходится – 5-6т.
Различия в урожаях в конечном итоге определяются тем, достигал ли и как скоро достигал посев в своем развитии оптимального для данных условий состояния, а также как долго он функционировал в этом состоянии.
Таким образом, высокие урожаи обеспечиваются определенным ходом фотосинтетической деятельности растений в посевах. Оптимальный ход нарастания площади листьев и биомассы может быть определен для каждой культуры и сорта в конкретных условиях выращивания.
Факторы, лимитирующие фотосинтез
Почему растения в посеве не достигают оптимального уровня показателей фотосинтетической деятельности или существенно отклоняются от них в отдельные периоды вегетации? Какие элементы фотосинтетической деятельности подвержены наибольшему влиянию различных факторов?
Как следует из формулы У=ФП*ЧПФ, урожай биомассы (а также хозяйственно ценной урожая) прямо зависит от этих показателей. При улучшении условий жизни растений (оптимизация режима питания и влагообеспеченности) обычно усиливаются ростовые процессы, увеличивается площадь листьев. В этом случае листья сильнее затеняют друг друга, поэтому ЧПФ снижаются.
Большинство культурных растений относятся к типу С3, у которых фотосинтез усиливается с ростом концентрации СО2 при насыщающей интенсивности освещения.
К растениям типа С4 относятся хлеба 2 группы – кукуруза, сорго, просо, рис и сахарный тростник. У них не наблюдается светового насыщения и усиленного фотодыхания, а компенсационная точка по СО2 необычайно низка. ЧПФ у С4-растений выше, чем у С3 растений, особенно при повышенной площади листьев. Сорта детерминантного типа роста (ультраскороспелые сорта сои) обычно характеризуются более высокой ЧПФ. Имеют значение также форма, направление и анатомическое строение листьев. У сортов люпина узколистного ЧПФ обычно выше, чем у сортов других видов люпина.
Считается, что у зерновых культур фотосинтез посева осуществляется лучше, если верхние листья направлены под острым углом к стеблю. В процессе селекционного совершенствования сахарной свеклы распластанная по поверхности почвы розетка листьев постепенно превращалась в воронкообразную, что определило способность современных сортов создавать высокопродуктивные посевы.
Большое значение имеет аттрагирующая способность генеративных и запасающих органов растений, благодаря которой пластические вещества из листьев активно перемещаются в них.
Для скороспелых форм ФП должен составлять – 1,5-2млн м?*дни/га, для среднеспелых – 2,5-3 и для позднеспелых – 3-5млн м?*дни/га.
В пределах одного типа скороспелости увеличение ФП может происходить путем ускорения темпов роста площади листьев и увеличения ее максимального значения. Густота посевов – один из факторов, сильно влияющих на эти показатели. Однако чрезмерное разрастание листьев при усилении их затенения может стать отрицательным фактором для формирования репродуктивных органов.
При оптимально влагообеспеченности фактором, ограничивающим ростовые процессы, становится уровень минерального питания. Для получения высоких урожаев необходима высокая обеспеченность элементами минерального питания, в первую очередь азотом. Так, растения пшеницы при урожайности зерна – 5т/га выносят из почвы – 170-200кг азота/га. Однако высокие нормы азота при достаточном количестве влаги усиливают вегетативный рост растений и разрастание листовой поверхности сверх оптимальных размеров, растения полегают, урожай снижается.
Основное средство повышения эффективности использования ценозом влаги и удобрения на фотосинтез и повышение урожайности – генетическое преобразование растений, создание сортов, у которых рост вегетативных органов ограничен, а генеративных – значительно выше, чем у старых сортов. Поэтому одно из направлений современной селекции – создание сортов, реагирующих на улучшение условий выращивания увеличением хозяйственно ценной части урожая.
В онтогенезе растений от всходов до начала созревания выделяют 4 биологически обоснованных периода:
- Всходы – начало цветения
- Цветение и образование плодов
- Рост плодов
- Налив семян
Основные выходные показатели первого периода – площадь листьев и ФП; второго – максимальная за вегетацию площадь листьев, ФП этого периода и количество образовавшихся плодов в расчете на единицу площади (на 1м?). Число образовавшихся плодов прямо коррелирует с ФП первого периода и особенно тесно – с ФП второго.
В третий период (рост плодов) площадь листьев постепенно начинает уменьшаться, но в среднем сохраняется на высоком уровне; биомасса продолжает интенсивно нарастать за счет роста плодов, которые к концу периода достигают максимальной величины. От числа сохранившихся к концу третьего периода плодов и семян зависят показатели фотосинтеза и поступление в растения азота в 4 период, когда происходит налив семян. Посев как фотосинтезирующая система наиболее активно функционирует в течение 2 и 3 периодов. За это время, равное по продолжительности первому периоду, накапливается – 60-70% биомассы и такое же количество азота от максимальных за вегетацию величин.
Таким образом, эффективность работы каждого последующего периода зависит не только от агрометеорологических условий этого периода, но и от результатов функционирования посева в предыдущий период. Конечные показатели развития посева – урожай семян и его качество – зависят от последовательных состояний посева в процессе роста и развития растений.
Органическое вещество, составляющее биомассу растений, образуется в процессе фотосинтеза из углекислого газа и воды за счет энергии поглощенной растениями ФАР. Связанная в урожае энергия ФАР является, таким образом, энергетическим выражением продуктивности фитоценоза.
Уже первые исследования показали, что между интенсивностью фотосинтеза единицы площади листа в единицу времени и накоплением биомассы растения отсутствует прямая зависимость (Тагеева, 1931; Катунский, 1941). В балансовом уравнении Иванова (1941) общая продуктивность фотосинтеза рассматривается как функция средней за учетный период интенсивности фотосинтеза величины фотосинтезирующей поверхности и рабочего времени. Уравнение учитывает также интенсивность дыхания и объем дышащей массы растения.
С развитием работ по проблеме "Фотосинтез и урожай" возникло представление о фотосинтетической деятельности растений как сложной совокупности процессов, в основе которых лежит поглощение растениями в фитоценозах энергии ФАР, и использование ее в процессе фотосинтеза на формирование урожаев (Ничипорович, 1956, 1966, 1966а, 1977).
Фотосинтетическая деятельность растений характеризуется интенсивностью фотосинтеза листьев и качественным составом образующихся в них биопродуктов, ходом роста вегетативных органов и листовой поверхности, накоплением биомассы растений, распределением продуктов фотосинтеза между вегетативными и репродуктивными органами и др.
Фотосинтетическая деятельность растений находится под постоянным влиянием состояния и динамики внешней среды. В отличие от одиночно стоящего растения на фотосинтетическую деятельность растений в ценозах оказывает влияние ценотическое взаимодействие растений, проявляющееся в конкуренции растений за условия жизни.
Как процесс формирования урожая фотосинтетическая деятельность растений подчиняется определенным законам и поддается количественному и качественному описанию, моделированию и управлению.
В основе работ по программированному получению урожаев лежит уравнение урожая, предложенное Ничипоровичем (1956). Уравнение дает количественное выражение зависимости биологических урожаев от размеров и работы листового аппарата с учетом качественной направленности процесса:
где Убиол — общий вес сухой биомассы растения; Фсо2 — интенсивность фотосинтеза (количество усвоенной СO2 в g/m2.день); Кэф— коэффициент эффективности фотосинтеза (отношение фактически накопленной сухой массы урожая к количеству усвоенной СO2); Л—площадь листьев (m2/ha); и — число дней продукционного периода.
Общий биологический урожай (Убиол) представляет собой сумму суточных приростов сухой массы растения. Зависимость хозяйственного урожая (зерно, плоды и др.) от биологического урожая представляется уравнением
где Кхоз — коэффициент хозяйственной эффективности фотосинтеза, характеризующий долю сухого вещества Ухоз от массы Убиол.
Для характеристики фотосинтетической деятельности растений наряду с размерами площади листьев используется показатель фотосинтетического потенциала (ФП). Он выражается в m2. днях и представляет собой сумму ежедневных площадей листьев растения или ценоза за вегетационный период или за какую-то его часть (ФП= ΣЛ.п). Показатель ФП является одним из основных при программировании урожая. Выражение ФСO2 х Kэф характеризует чистую продуктивность фотосинтеза (ЧПФ, или ,,нетто-ассимиляцию“), т. е. весовое количество суточного прироста сухого вещества растения в расчете на 1 m2 площади листьев. С учетом указанных преобразований уравнение (5) получит следующий вид:
Суточный прирост общей сухой массы биологического урожая определяется как произведение
На основе данных по фотосинтетической деятельности (Л, ФП, ЧПФ, С) с учетом факторов обеспечения (вода, минеральное питание и др.) определяется предполагаемое или планируемое нарастание биомассы по фазам вегетации на протяжении вегетационного периода для получения заданной величины Убиол с оптимальной структурой урожая (Кхоз).
Наивысшие урожаи могут быть получены при следующих оптимальных условиях (Ничипорович, 1956):
а) быстром развитии и сохранении в течение возможно длительного времени в активном состоянии большей площади листьев (Л);
б) наиболее высоких величинах интенсивности и коэффициентах эффективности фотосинтеза;
в) наиболее высокой чистой продуктивности фотосинтеза (ЧПФ) и высоких суточных приростах (С) сухого вещества;
г) наилучшем распределении и использовании образуемых веществ на формирование хозяйственной части урожая (Кхоз);
д) наиболее полном использовании потенциального периода вегетации (n).
Таблица 1
Зависимость величины урожая винограда (kg) от площади листьев (т2)
1 kg сахара ягод
Kozina (цит. по Стоеву, 1973)
Couvillon, Nakayama, 1970
Liuni (цит. по Стоеву, 1973)
Тодоров, Занков, 1964
Мельник, Анисимова, 1953
Для программирования урожая винограда представляет интерес показатель "продуктивности листьев", характеризующий количество сырого урожая или сахара гроздей 1 , произведенное в расчете на единицу площади листьев.
Следует отметить, что простой пересчет сырой массы или валового сахара гроздей на листовую площадь растения характеризует фактически сложившееся соотношение между площадью листьев и гроздями, но не потенциальную продуктивность листьев (ПЛ). Для получения характеристики потенциальной ПЛ необходимо, чтобы площадь листьев по отношению к гроздям находилась в оптимуме или ограничивала бы рост гроздей. Это достигается в опытах с различной нагрузкой побегами или путем частичного удаления листьев с побега при одновременном их кольцевании (Амирджанов, 1963).
В табл. 1 показаны полученные разными исследователями данные по продуктивности листьев винограда. Для возможности сопоставления они приведены нами к одной характеристике: площади листьев, необходимой для получения единицы урожая (1 kg сырой массы гроздей или 1 kg сахара гроздей).
Величина ПЛ винограда варьирует в широких пределах; от 3 до 15 m2/kg сахара и от 0,7 до 2,0 m2 /kg сырого урожая гроздей, что, очевидно, в большей степени вызвано разными методами расчета, чем фактической потенциальной продуктивностью листьев. Если с известным допущением усреднить соответствующие крайние значения, то необходимая площадь листьев составит для получения 1 kg сырого урожая ягод — 1—1,5 m2; для получения 1 kg сахара гроздей — 6—9 m2.
Эти величины практически идентичны для ягод с сахаристостью сока 17—18% при содержании сока в ягодах 80%.
Таким образом, по самой приближенной оценке при благоприятных условиях для получения 100 cwt гроздей при сахаристости сока ягод 17—18 % необходимо иметь на гектаре - 10—15 тыс. m2 площади листьев. Количество побегов, соответствующее этой площади листьев и необходимое для получения заданного урожая, будет зависеть от фитометрических характеристик побега и показателей его плодоносности: коэффициента плодоношения 2 и средней массы грозди или, иначе говоря, от продуктивности одного побега.
Как показывают исследования, урожаи наиболее тесно коррелируют с размерами листовой поверхности. Следовательно, для получения планируемого урожая винограда необходимо иметь на гектаре такое количество нормально развитых побегов, ход роста которых обеспечил бы получение расчетной величины ФГТ. Это обстоятельство указывает на важность изучения закономерностей роста побегов винограда в зависимости от нагрузки побегами и гроздями, длины обрезки, формировки, фона питания и др. с целью установления "оптимального" хода роста.
При оптимальных условиях произрастания растения должны не только осуществлять большую фотосинтетическую работу, но и эффективно использовать продукты фотосинтеза на формирование хозяйственной части урожая— гроздей.
У винограда структура биологического урожая определяется как внешними факторами, так и природой самого растения. К числу "внутренних" факторов относятся такие, как сила роста куста, плодоносность побегов, величина грозди, нагрузка побегами и гроздями и др. У растений с одинаковой вегетативной массой величина Кхоз будет тем больше, чем выше плодоносность побегов. При одинаковой плодоносности уменьшение вегетативного прироста повлечет за собой повышение показателя Кхоз.
Величина Кхоз у винограда изменяется в больших пределах. У сорта Алиготе в зависимости от площади питания она изменялась от 0,45 до 0,57; у сорта Карабурну — от 0,60 до 0,67 (Плакида, 1967). У сорта Болгар (синоним Карабурну) в зависимости от высоты штамба величина Кхоз, изменялась от 0,48 до 0,71 (Панделиев, 1976). В опытах Стоева и Добревой (1976) у сорта Ркацители в приземной культуре Кхоз составил 0,45, а при штамбовой культуре — 0,73 при близких значениях Убиол (1,34 и 1,38 kg на куст).
1 Поскольку процентное содержание сахара в соке ягод в разных случаях неодинаково, правильнее характеризовать этот показатель не по сырому урожаю, а по количеству сахара гроздей.
2 Коэффициент плодоношения (Кпл) — среднее число гроздей на один развившийся побег.
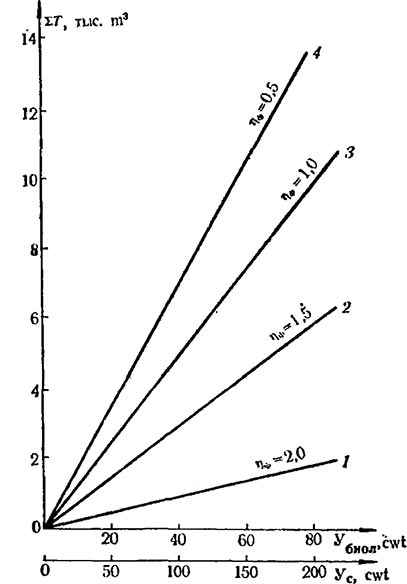
По нашим данным у сорта Шабаш в зависимости от густоты посадки и формировки кустов величина Кхоз изменялась от 0,33 до 0,46; у сорта Рислинг рейнский — от 0,33 до 0,52. У сортов Пино гри, Мускат белый и Хиндогны в среднем за пять лет величина Кхоз составила соответственно 0,30, 0,47 и 0,36.
Задача состоит в том, чтобы при помощи различных агротехнических приемов находить для каждого отдельного случая оптимальную величину Кхоз, т. е. добиваться наилучшего перераспределения ассимилятов на формирование хозяйственного урожая. Вместе с тем величину Кхоз у винограда нельзя увеличивать беспредельно. Превышение Кхоз некоторого оптимального для конкретного случая уровня практически будет означать перегрузку кустов гроздями, и это может служить причиной ухудшения качества ягод и ослабления силы кустов. С точки зрения создания высокопродуктивных насаждений с оптимальной структурой кустов важным представляется подбор сортов, сочетающих высокие показатели плодоносности с относительно умеренным ростом побегов.
Получение планируемых урожаев винограда требует знания механизма взаимодействия показателей фотосинтетической деятельности. Пример такого взаимодействия показан на рис. 1.
Рис. 1. Графическое представление механизма фотосинтетической деятельности винограда в насаждении
1 — сухая биомасса (У); 2 — потенциальный хозяйственный урожай (Ухоз); 3 — хозяйственный коэффициент (Кхоз); 4 — чистая продуктивность фотосинтеза (ЧПФ); 5 — реальный хозяйственный урожай (Ухоз 2)
С увеличением на гектаре площади листьев (ФП) растет величина Убиол, но не пропорционально, поскольку с ростом площади листьев снижается ЧПФ. По мере роста площади листьев (Л) повышается также величина Ухоз. Максимальные уровни Ухоз при заданных Л характеризуются линией регрессии Ухоз 1, которая получена при условии, что величина Кхоз оставалась постоянной (0,45 для сорта Шабаш). Однако по мере роста Л снижается Kхоз, и в результате реальные уровни хозяйственного урожая — Ухоз 2 отклоняются от максимальных. По мере увеличения Л насаждения разрыв между максимальными (Ухоз 1) и реальными (Ухоз 2) урожаями гроздей увеличивается; возникает как бы „эффект ножниц“. Уменьшение Кхоз в насаждениях с густой посадкой (имеющих высокие показатели Л) обусловлено снижением плодоносности побегов: Кпл и средней массы грозди.
Для разбираемого случая зависимость Убиол (cwt/ha) от ФП (млн.m2. дней) имеет линейный характер и описывается уравнением
Связь между Ухоз (cwt сухого вещества) и площадью листьев (тыс. m2) описывается квадратичным уравнением
Аналогичный характер связи выявлен Ординовой (1968) между сырым урожаем гроздей и густотой посадки виноградника. Следовательно, одной из задач оптимизации продукционного процесса винограда для получения планируемых урожаев является отыскание оптимальных размеров площади листьев насаждения (количества побегов и соответствующего их размещения в пространстве), обеспечивающих получение максимального хозяйственного урожая без снижения его качества. Планируемый урожай должен быть обеспечен водой и минеральным питанием.

Наибольший прирост листовой поверхности до фазы колошения озимой пшеницы составил по кулисно-мульчирующему пару - 12,49 тыс.м2/га, на чистом пару прирост составил 12,01 тыс.м2/га. По занятым парам наиболее интенсивно нарастали растения озимой пшеницы, посеянной по занятому донником пару - прирост составил 12,44 тыс.м/га. По данному предшественнику наблюдается и интенсивное снижение площади листовой поверхности к фазе молочного состояния зерна - на 22,38 тыс.м2/га. Менее интенсивное снижение наблюдается по пару, занятому эспарцетом и по кулисно-мульчирующему пару - на 21,66 и 20,17 тыс.м /га соответственно. К моменту молочного состояния зерна озимой пшеницы наибольшая площадь листовой поверхности растений была по кулисно-мульчирующему пару - 17,13 тыс.м2/га, что в 1,17 раз превышает показатели по чистому пару и в 1,25-1,27 раз больше площади листьев озимой пшеницы по занятым парам.
На образование сухого вещества большое влияние оказывает солнечная энергия, причем зависимость хозяйственного урожая различных культур от солнечной энергии варьирует соответственно тому, какую часть биомассы он составляет. У зерновых культур тесной зависимости не отмечено, но известно, что определенному урожаю зерна отвечает определенный уровень общего сухого вещества биомассы.
С динамикой увеличения площади листовой поверхности тесно связаны показатели накопления сухого вещества по фазам вегетации (табл. 3.4.2).

Так, наибольшее количество сухого вещества сформировали растения озимой пшеницы в фазу выхода в трубку по кулисно-мульчирующему пару - 289,5 г/м2, что на 8,3 г/м больше показателя по чистому пару и на 33,1-37,0 г/м2 больше, чем по занятым парам. К фазе колошения растения озимой пшеницы интенсивно формировали свою надземную массу - увеличение количества сухого вещества по сравнению с фазой выход в трубку составило в 1,75-1,82 раза, а к фазе полной спелости - в 2,94-3,11 раз.
Наибольшее количество сухого вещества к фазе полной спелости сформировала озимая пшеница по кулисно-мульчирующему пару - 873,4 г/м2, что на 47,3-89,2 г/м2 выше показателей чистого и занятых паров. Таким образом, чем большую площадь листьев формируют растения, тем больше они накапливают сухого вещества, так как листовой аппарат растений озимой пшеницы работает более длительный период для создания сухого вещества.
В умеренном поясе основным лимитирующим фактором роста служит энергия солнечных лучей. С ней связано и влияние температуры, зависимость формирования урожая от которой характеризуется: чем больше тепла в пределах оптимальных колебаний для данного вида получают растения, тем лучше они растут. Следует подчеркнуть, что это относится к температурам, при которых активность роста максимальная. Co снижением или, наоборот, повышением температуры относительно оптимальных величин активность роста ослабевает, а при предельных ее значениях (минимальная, максимальная) совсем прекращается. Это является одной из причин, по которым на вариантах с кулисно-мульчирующими парами озимая пшеница имеет большую площадь листьев и накапливает больше органического вещества на единицу площади.
Для комплексной и правильной оценки фотосинтетической продуктивности листового аппарата растений озимой пшеницы используется показатель, который объединяет площадь листовой поверхности растений и продолжительность работы листового аппарата -это фотосинтетический потенциал. Как отмечает А.А. Ничипорович, фотосинтетический потенциал характеризует производительную способность растений как за разные периоды вегетации, так и в целом за вегетацию. При анализе фотосинтетического потенциала растений озимой пшеницы нами установлено, что наилучшие условия для роста и развития созданы по кулисно-мульчирующему пару (табл. 3.4.3).
В межфазный период кущение-выход в трубку фотосинтетический потенциал растений озимой пшеницы по чистому пару составил 704,2 тыс. м2/га*сутки, по занятым донником и эспарцетом парам - на 35,9-42,0 тыс. м2/га*сутки меньше вследствие меньшей площади листовой поверхности растений.


Читайте также: