
Объясните роль полиплоидии в создании новых сортов растений
Добавил пользователь Дмитрий К. Обновлено: 18.09.2024
Чрезвычайно ценным источником изменчивости для селекции растений служит полиплоидия.
Увеличение числа наборов хромосом у культурных растений сыграло выдающуюся роль в эволюции видов и селекции. Народная селекция, не зная самого явления полиплоидии, давно использовала ее как источник изменчивости в создании ряда таких ценнейших культур, как пшеница, овес, хлопчатник, картофель, а также в цветоводстве.
Изучение полиплоидии позволило овладеть этим источником изменчивости растений. Успехи теоретических исследований не замедлили сказаться на получении искусственных полиплоидов у сельскохозяйственных культур.
Число искусственно созданных полиплоидов с каждым годом прогрессивно нарастает. В настоящее время только у ржи получено несколько десятков тетраплоидов.
Применение колхицина ускорило получение полиплоидов. При этом успех зависит от способа обработки тканей, вида растения, стадии воздействия. Раствором колхицина разной концентрации обрабатывают семена, проростку точки роста взрослого растения, а также воздействуют через корневую систему.
Автополиплоидия и аллополиплойдия имеют разную ценность в селекционной работе. Удвоение числа хромосом от диплоидного к тетраплоидному часто сопровождается увеличением размера или ускорением темпа деления клеток, что ведет к изменению размеров растения и его органов, размера и веса семян, химического состава, а также к повышению устойчивости к заболеваниям и т. д. Так, например, вес 1000 зерен тетраплоидной ржи сорта Стальной составляет 45—50 г, а диплоидной того же сорта — 28—30 г. Вес 1000 зерен тетраплоидной гречихи, полученной В. В. Сахаровым, составляет 23—35 г, диплоидной — 16—29 г, и т. д. В настоящее время советскими генетиками получены полиплоиды у ряда сельскохозяйственных культур: сахарной свеклы, проса, кок-сагыза, опийного мака, льна, перечной мяты, редиса, кукурузы, земляники и других растений.
Вместе с тем автополиплоиды часто характеризуются снижением фертильности. Хотя каждое семя у полиплоида крупнее, чем у исходной формы, но количество семян на растении может быть меньше. Причины этого кроются в основном в нарушении мейоза. Однако это препятствие преодолимо. Полиплоид не является готовым сортом — он требует дальнейшей тщательной селекции, а возможно, и скрещивания с другими полиплоидами, а также последующего отбора гибридных форм.
В селекции широко используются как естественные аллополиплоиды, так и экспериментально полученные от скрещивания разных видов и родов. Полиплоидия особенно эффективна при низкой урожайности стандартных сортов. При высокой урожайности сортов она хотя и дает прибавку урожая, но не столь разительную. Вместе с тем полиплоидия (автополиплоидия) часто сопровождается изменением хозяйственно ценных признаков. Например, автополиплоиды ржи имеют более прочную солому (что препятствует полегаемости), но при этом пониженную фертильность. Однако снижение фертильности автополиплоидов может быть преодолено либо путем селекции, либо путем создания искусственной популяции, составленной из линий автополиплоидов разного происхождения. Такие самовоспроизводящиеся, синтетические, популяции автополиплоидов могут сохраняться в течение нескольких поколений без заметного снижения урожайности. Следует подчеркнуть, что автополиплоидия вызывает больший эффект у перекрестноопылителей, чем у самоопыляющихся растений.
Современные цитогенетические исследования открыли новый путь, раскрывающий широкие перспективы для селекции, — использование в сельскохозяйственном производстве триплоидных форм. Триплоидные растения обычно являются или стерильными, или в большинстве своем имеют низкую фертильность. Однако в ряде случаев триплоидные растения отличаются повышенной мощностью и урожайностью по вегетативной массе, по весу корнеплода, например, у сахарной свеклы.
Японскими генетиками была разработана система скрещивания тетраплоидного (2n = 44) и диплоидного (2n = 22) арбуза. Гибриды между ними являются триплоидными, а поэтому бессемянными; они имеют очень крупные плоды и большую устойчивость к заболеваниям.
Однако важно подчеркнуть, что выращивание триплоидных растений, в частности арбузов, требует определенной системы семеноводства: во-первых, необходимо получение и селекция тетраплоидных линий, которые должны поддерживаться в чистоте; во-вторых, для получения высокоурожайных триплоидов требуется подбирать линии автотетраплоидов и диплоидов из разных сортов с высокими комбинационными свойствами, дающими при скрещивании гетерозис; в-третьих, триплоидные семена в плодах образуются чаще в результате такого скрещивания, когда тетраплоид является женским растением, а диплоид — мужским. Реципрокная комбинация скрещивания не дает семян. Это еще раз показывает, что совместимость геномов с цитоплазмой и степень нарушения мейоза зависят от направления скрещивания.
Пыльца тетраплоидов оказывается менее жизнеспособной, чем яйцеклетки.
Для того чтобы выяснить, с какой вероятностью у арбуза могут встречаться плоды с семенами, следует познакомиться с ходом мейоза у триплоидных растений. У триплоида в первом делении мейоза образуются не биваленты, а триваленты, при расхождении которых анафазе две хромосомы могут отойти к одному полюсу, а третья — противоположному. В результате такого деления образуются гаметы с числом хромосом от 11 до 22; частота появления каждого класса гамет вычисляется по уравнению (1 + 1) 11 . Фертильными могут быть только два сорта гамет — с 11 и с 22 хромосомами, гаметы с промежуточным числом хромосом окажутся стерильными. Следовательно, фертильных гамет будет 0,5 11 X 2, т. е. 0,1%, а стерильных остальные 99,9%. На основании такого расчета в потомстве триплоида следует ожидать, как правило, бессемянные плоды. Плод триплоидного арбуза развивается партенокарпически в силу стимуляции его веществами, привносимыми пыльцой, попадающей на рыльце пестика. Если изолировать цветки триплоидного арбуза от пыльцы, то завязь не развивается.
Мы подробно остановились на примере получения триплоидного арбуза для того, чтобы иллюстрировать разные аспекты использования полиплоидии в практических целях.
Таким образом, получение полиплоидов дает в руки селекционера формы, измененные в качественном и количественном отношениях, Однако, следует иметь в виду, что искусственно полученные полиплоиды являются лишь исходным материалом для отбора, с ними необходимо вести дальнейшую селекционную работу, приспосабливая их к требованиям производства, климату и агротехнике.
При изучении генетики полиплоидов в практических целях крайне важно для каждого вида растений установить 1) совместимость геномов между собой и цитоплазмой, 2) изменение характера Доминирования и взаимодействия генов при умножении числа геномов.
Действие мутантных генов в гетерозиготном состоянии, превышающих и понижающих жизнеспособность и продуктивность растений.
При получении полиплоидов в селекционных целях необходимо иметь в виду следующее:
- полиплоиды перспективнее получать от сортов, имеющих высокую продуктивность в условиях данного района возделывания, и от гибридов между специально подобранными сортами разного происхождения и инбредных линий, обладающих высокими комбинационными свойствами;
- полиплоидия не для всех видов растений может быть успешной в хозяйственных целях;
- методика экспериментального получения полиплоидов различна для разных видов.
Искусственно создаваемая полиплоидия дает в руки селекционера средство, позволяющее сокращать время эволюции растения в культуре.
В заключение рассмотрения источников изменчивости в селекции следует подытожить ряд общих положений о ее значении в эволюции культурных растений. Первичным источником эволюции культурных растений являются мутации всех типов, которые при гибридизации и в процессе отбора комбинируются в системы генотипов. Поэтому эволюция культурных растений протекает на основе отобранных систем генотипов в условиях возделывания.
В ряде случаев установлено, что вновь получаемые мутанты оказываются не новыми для данной культуры, т. е. в том или ином виде они уже известны в мировой коллекции сортов. Под руководством Г. Штуббе в ГДР ведутся широкие исследования по сопоставлению спонтанных и экспериментально полученных мутантов у ячменя, томатов, сои с имеющимися в культуре разновидностями. И оказывается, что, например, 170 известных мутантов для ячменя укладываются в существующую систему признаков 192 ясно различимых разновидностей. Сходная картина установлена для томатов, львиного зева и других растений. Отсюда Г. Штуббе делает очень важный вывод о том, что все естественное многообразие форм, например, ячменя, может быть воспроизведено путем экспериментального получения мутаций с последующей комбинацией их при скрещиваниях и отборе. Автор считает, что уже сейчас экспериментально созданное многообразие мутантов и известное природное многообразие форм ячменя, собранных в мировой коллекции, ничем существенным не отличаются. И действительно, ряд экспериментальных мутаций генетически идентифицируется при локализации с таковыми у существующих разновидностей ячменя, томатов, сои и львиного зева.
Из этого видно, что закон гомологических рядов Н. И. Вавилова получает реальное воплощение в анализе эволюции культурных растений.
Накопленный в процессе эволюции фонд мутаций культурных растений еще не означает, что для получения новых мутаций уже достигнут предел. В процессе селекции культурных растений многие аллели различных генов и их комбинаций, наверное, не были еще оценены искусственным отбором, так как они редко спонтанно мутируют. Поэтому индуцирование мутаций остается главным источником наследственной изменчивости для отбора в селекции культурных растений.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
В 1892 году русский ботаник И.И. Герасимов исследовал влияние температуры на клетки зеленой водоросли спирогиры и обнаружил удивительное явление - изменение числа ядер в клетке. После воздействия низкой температурой или снотворным (хлороформом и хлоралгидратом) он наблюдал появление клеток без ядер, а также с двумя ядрами. Первые вскоре погибали, а клетки с двумя ядрами успешно делились. При подсчете хромосом оказалось, что их вдвое больше, чем в обычных клетках. Так было открыто наследственное изменение, связанное с мутацией генотипа, т.е. всего набора хромосом в клетке. Оно получило название полиплоидии, а организмы с увеличенным числом хромосом - полииплоидов.
В природе хорошо отлажены механизмы, обеспечивающие сохранение постоянства генетического материала. Каждая материнская клетка при делении на две дочерние строго распределяет наследственное вещество поровну. При половом размножении новый организм образуется в результате слияния мужской и женской гаметы. Чтоб сохранилось постоянство хромосом у родителей и потомства, каждая гамета должна содержать половину числа хромосом обычной клетки. И в самом деле, происходит уменьшение в два раза числа хромосом, или, как назвали ученые редукционное деление клетки, при котором в каждую гамету попадает только одна из двух гомологичных хромосом. Итак, гамета содержит гаплоидный набор хромосом - т.е. по одной от каждой гомологичной пары. Все соматические клетки диплоидны.
Полиплоидия (от греч. polyploos - многократный и eidos - вид), кратное увеличение числа наборов хромосом в клетках организмов. В естественных условиях возникает под влиянием различной природных факторов (резкая смена температур, действие ионизирующей радиации и др.), а также при естественной гибридизации. В основе механизма действия этих факторов лежит не расхождение
хромосом в митозе (т. н. митотическая полиплоидия, напри-мер, у голосеменных) или в мейозе (мейотическая полиплоидия). При поли-плоидии нормальный диплоидный набор хромосом (2 n), свойственный соматическим клеткам большинства организмов, увеличивается кратно гаплоидному набору (n), свойственному половым клеткам.
Полиплоидизация - перспективный метод в селекции лесных пород, ведущий к улучшению существующих и созданию новых, более совершенных форм. Искусственные триплоиды тополя, берёзы, ольхи и других древесных пород, получаемые скрещиванием диплоидных и тетраплоидных форм и отличающиеся быстрым ростом, прочной и устойчивой к гнилям древесиной, декоративностью, высоко ценятся в лесоразведении и озеленении.
Полиплоидия – это естественный природный процесс видо- и формообразования, ведущий к улучшению существующих и возникновению новых более совершенных форм растений. Изучение некоторых разновидностей и сортов культивируемых древесных растений показало, что по своей генетической природе они являются аллополиплоидамми древних гибридных видов. Между количеством хромосом и свойствами полиплоидных форм не всегда наблюдается прямая связь. Часто диплоидные особи оказываются значительно лучше полиплоидов, полученных от них. Например, тетраплоидные формы березы, тополей, вязов, ольхи и робинии лжеакации растут медленнее диплоидных, а триплоидные особи оказались более быстрорастущими. Существует известный оптимум числа хромосом, обеспечивающий наиболее эффективный рост и накопление практически ценных для селекции качеств. У древесных растений оптимальным уровнем полиплоидии считается триплоидный. Например, триплоидная исполинская осина (Populus tremula var gigas) характеризуется быстрым ростом, увеличенным размером клеток и органов, устойчивостью к грибным заболеваниям. Разработаны методы массового искусственного получения триплоидной осины. Растения с очень большим числом хромосом нередко имеют пониженную жизнеспособность.
Больше полиплоидов среди лиственных пород (особенно в семействах ивовых, берёзовых, розовых, бобовых и др.), меньше - среди хвойных. Около трети всех видов покрытосеменных возникло за счёт полиплоидии (среди голосеменных только 4,5% видов - полиплоиды). Полиплоидия ведёт к образованию гетерозисных форм, характеризующихся мощностью развития, усиленным и быстрым ростом, накоплением в тканях ценных химических соединений и т. д. Однако повышение плоидности, ведущее к усилению мощности растений, имеет определенные границы. Так, у цитрусовых деревьев (апельсин, мандарин, лимон) искусственная полиплоидия вызывает карликовость.
Особенно большое значение в селекции растений, в том числе и древесных, имеет восстановление фертильности (плодовитости) аллополиплоидов.
Фертильность определяется порядком прохождения мейоза. У тетраплоидных особей в клетках растения существуют два набора гомологичных хромосом, которые в мейозе группируются по четыре хромосомы. Четыре гомологичные хромосомы могут расходиться к противоположным полюсам следующим образом: 2 и 2, 2 и 3, 3 и 1. Дочерние клетки в результате мейоза могут получить неполный или избыточный набор хромосом. Гаметы с несбалансированным набором хромосом, как правило, погибают. Такому стерильному дереву с нарушенным процессом образования гамет можно вернуть плодовитость (фертильность) и вмешательством в процесс формирования кариотипа зародыша семян. Это имеет большое значение при отдаленной гибридизации для восстановления фертильности в последующих поколениях ценных гибридов.
Существует правило: если диплоид фертильный, то образованный от него тетраплоид может быть стерильным; если же диплоид стерильный, то производный тетраплоид может быть фертильным. Наряду с этим замечено, что если в первом гибридном поколении (F1) диплоидный гибрид стерилен или образует варьирующее потомство, то при удвоении числа хромосом он может стать фертильным и хорошо размножаться, так как в этом случае все члены одного набора хромосом полностью соответствуют своим двойникам в другом наборе хромосом. Например, полученный гибрид с диплоидным набором хромосом имеет негомологичные хромосомы вследствие скрещивания отдаленных в систематическом отношении видов. В результате хромосомы не образуют во время мейоза необходимых гомологичных пар и не распределяются равномерно по полюсам. Вместо этого они могут мигрировать к полюсам в сочетании: 2 и 0, 1 и 1 , 0 и 2. В каждом случае яйцеклетка или пыльцевое зерно получает несбалансированные наборы хромосом и погибает. Однако если число хромосом гибрида удваивается и образующаяся тетраплоидная особь имеет два полных набора хромосом (по 2п в каждом), получаются гаметы с гомологичными наборами хромосом. В мейозе хромосомы образуют пары и равномерно распределяются в дочерние клетки.
Анализируя все вышесказанное можно сделать вывод: при полиплоидии в ядрах клеток растений происходит увеличение числа хромосом в кратном отношении к гаплоидному (п), при этом с увеличением числа хромосом на один набор 2 + 1 = 3 появляются триплоидные растения, при 2 + 2 = 4 – тетраплоиды и т.д. Увеличение числа хромосом может быть неограниченно большим, но наиболее эффективны небольшие их величины.
Полиплоиды могут возникать в естественных условиях под влиянием природных факторов: резких смен температур, космической радиации, гроз, бурь, ливней, града и т.п.
Полиплоидия может быть митотического (изменяется набор хромосом в соматических клетках) и мейотического (изменяется набор хромосом в половых клетках) происхождения, последняя бывает реже.
Все полиплоиды могут быть объединены в четыре группы:
1 автополиплоиды – организмы, имеющие увеличение гаплоидного набора более, чем в два раза хромосом одного и того же вида.
2 аллополиплоиды – особи, возникающие на основе объединения или умножения двух или нескольких целых геномов, принадлежащих разным видам или родам.
3 анеуплоиды – организмы, имеющие уменьшенное или увеличенное число хромосом некратное основному числу хромосом вида.
4 гаплоиды – организмы, имеющие в соматических клетках не увеличенное, а уменьшенное число хромосом.
Следовательно, в селекционной работе с древесными растениями полиплоидия приобретает значение, как метод восстановления фертильности
и преодоления явления несовместимости при отдаленной межвидовой гибридизации.
Существует ряд методов получения полиплоидов. Так методы получения аллоплоидов можно разделить на две группы: опыление предуцированными гаметами и индуцирование мутагенами. При нормально протекающем мейозе хромосомное число в клетках уменьшается с 2п до 1п. Иногда редукционного деления не происходит, и образуются яйцеклетки или пыльцевые зерна с 2п хромосомами. Если пыльцевое зерно с 2п хромосомами оплодотворит яйцеклетку с гаплоидным набором хромосом, образуется триплоидный эмбрион. Таким путем получают тетраплоидную осину, опыляя крупными триплоидными пыльцевыми зернами женские цветки диплоидных растений. Крупную триплоидную пыльцу отделяют от диплоидной, просеивая через тонкую ткань.
Индуцирование полиплоидов в свою очередь производится тремя способами:
1 Воздействие нагреванием, высушиванием или холодом. Удвоение соматических хромосом таким путем происходит довольно часто (в 1% или более делящихся клеток). Образующиеся клетки с тетраплоидным набором хромосом обрастают вокруг тканью и растение в целом остается диплоидным.
Получение триплоидов из семян. Триплоидные растения древесных видов характеризуются быстрым ростом. Поэтому получение тетраплоидов для последующего перевода их в триплоиды скрещиванием с диплоидами становится распространенным направлением в селекции на гетерозис. Так, в северной Европе в больших количествах выращивают триплоидные деревья осины, которые получают в результате скрещивания женских клонов тетраплоидных растений с одним из диплоидных мужских клонов. Эту работу, как правило, проводят в теплицах. Дополнительно к этому практикуют посадку одного тетраплоидного клона в центре естественных насаждений мужских диплоидных особей. На деревьях женского клона семена будут триплоидными.
Гаплоидные особи образуются в результате самопроизвольного деления неоплодотворенной яйцеклетки. Такие формы обнаружены, как у голосеменных, так и у покрытосеменных. Однако гаплоидные формы древесных пород в селекции не применяются, так как они не жизнеспособны.
Из неоплодотворенных яйцеклеток индуцируют развитие партеногенетических особей после опыления их заранее инактивированной пыльцой. Среду для культуры тканей обычно готовят из агара с добавлением веществ, стимулирующих рост: сахарозы, аминокислот, гормонов растений, минеральных веществ, экстрактов пыльников. Температурный режим и свет
– важнейшие факторы при культуре тканей растений. Пыльцевые зерна в культуру тканей берут в период, когда генеративное и вегетативное ядра начинают одновременно делиться. При этом имеется большая вероятность образования триплоидных эмбрионов, чем гаплоидных.
Такие методы успешно используются при работе с травянистыми растениями. Это дает основание полагать, что эти методы можно будет применять и при работе с древесными растениями.
Таким образом, можно сделать вывод, что изменение количества хромосом в кариотипе – важнейший источник формообразования исходного материала для селекции растений.
Полиплоидия и отдаленная гибридизация в селекции растений. Многие культурные растения полиплоидны, т. е. содержат более двух гаплоидных наборов хромосом. Среди полиплоидов оказываются многие основные продовольственные культуры: пшеница, картофель, овес. Поскольку некоторые полиплоиды обладают большой устойчивостью к действию неблагоприятных факторов и хорошей урожайностью, их использование в селекции оправданно.
Существуют методы, позволяющие экспериментально получать полиплоидные растения. За последние годы с их помощью созданы полиплоидные сорта ржи, гречихи, сахарной свеклы.
Отдаленная гибридизация, т. е. скрещивание растений, которые относятся к разным видам и даже родам, перспективна для создания совершенно новых форм растений. Однако гибриды первого поколения, как правило, бесплодны. Причина бесплодия заключается в нарушении конъюгации хромосом в мейозе. Полиплоидизация отдаленных гибридов приводит к восстановлению плодовитости благодаря нормализации мейотического процесса. Впервые отечественный генетик Г. Д. Карпеченко в 1924 г. на основе полиплоидии создал плодовитый капустно-редечный гибрид (рис. 50). Капуста и редька в диплоидном наборе имеют по 18 хромосом (2n=18).

Рис. 50. Процесс преодоления бесплодия межвидового гибрида на основе полиплоидизации (капустно-редечный гибрид)
Вследствие этого возникли нормальные условия для мейоза: хромосомы капусты и редьки соответственно конъюгировали между собой. Каждая гамета несла по одному гаплоидному набору редьки и капусты (9+9=18).
В зиготе вновь оказалось 36 хромосом; гибрид стал плодовитым.
Мягкая пшеница — природный полиплоид, состоящий из шести гаплоидных наборов хромосом родственных видов злаков. В процессе ее возникновения отдаленная гибридизация и полиплоидия играли важную роль.
Методом полиплоидизации отечественные селекционеры создали ранее не встречавшуюся в природе ржано-пшеничную форму — тритикале. Создание тритикале — нового вида зерновых, обладающего выдающимися качествами, — одно из крупнейших достижений селекции. Он был выведен благодаря объединению хромосомных комплексов двух различных родов — пшеницы и ржи. Тритикале по урожайности, питательной ценности и другим качествам превосходит обоих родителей. По устойчивости к неблагоприятным почвенноклиматическим условиям и наиболее опасным болезням она превосходит пшеницу, не уступая ржи.
В настоящее время генетики и селекционеры создают новые формы злаков, плодовых и других культур с использованием отдаленной гибридизации и полиплоидии.
Полиплоидия и отдаленная гибридизация у животных. Полиплоидия у животных в природе встречается редко. Однако у некоторых видов домашних животных она возможна. Известный отечественный ученый Б. Л. Астауров впервые создал полиплоидные формы тутового шелкопряда. Он пошел по пути, сходному с тем, который предложил Г. Д. Карпеченко. Используя отдаленную гибридизацию и полиплоидию, он создал совершенно новую форму тутового шелкопряда, объединившую в своем геноме хромосомы двух разных видов. Эта работа, несомненно, относится к числу блестящих достижений современной биологии.
Искусственный мутагенез и его значение в селекции. Вскоре после того, как было показано, что воздействие рентгеновскими лучами резко увеличивает темп мутационного процесса и вызывает массу новых мутаций, генетики приступили к разработке методов искусственного получения мутаций для целей селекции. В качестве мутагенов, т. е. вызывающих мутации агентов, в настоящее время широко используют различные виды ионизирующей радиации (рентгеновские лучи, гамма-лучи, тепловые и быстрые нейтроны, ультрафиолет) и специальные химические соединения.
В большинстве случаев мутации, возникающие у организмов под влиянием мутагенов, неблагоприятны для их носителей. Но наряду с мутациями, резко снижающими жизнеспособность, возникают и такие, которые могут представлять интерес для селекции.
Искусственный мутагенез, т. е. контролируемый человеком процесс возникновения мутаций, успешно применяется в селекции растений и микроорганизмов. Использование этого метода оказалось особенно эффективным применительно к разнообразным микроорганизмам: грибам, дрожжам, водорослям, бактериям. Микроорганизмы широко используются в пищевой промышленности, в производстве лекарств, биологически активных веществ, а также в производстве кормов для животных. Области их применения постоянно расширяются. С помощью искусственного мутагенеза, в частности, получены плесневые грибки, продуцирующие антибиотики в тысячи раз эффективнее, чем исходные формы. Значение антибиотиков общеизвестно: они сохраняют жизнь миллионам людей. Использование искусственного мутагенеза в селекции привело к созданию высокопродуктивных штаммов микроорганизмов — продуцентов витаминов, аминокислот, белков, которые активно используются в медицине и сельском хозяйстве. В будущем роль микроорганизмов в различных отраслях человеческой жизни неизбежно возрастет, что еще больше повысит значение селекции и генетики микроорганизмов.
Благодаря использованию мутагенов возникают мутантные формы растений, у которых сохранены ценные свойства формы или сорта и вместе с тем улучшены признаки, особенно интересующие селекционеров.
Мутантное растение лишь в редких случаях может быть сразу родоначальником сорта. В большинстве случаев такие растения служат материалом, который может быть использован для создания сортов путем гибридизации и отбора. Таким образом, искусственный мутагенез является важным и эффективным в арсенале методов, используемых в современной селекции.
В полиплоидия Это тип генетической мутации, заключающийся в добавлении полного набора (полных наборов) хромосом к ядру клетки, образующих гомологичные пары. Этот тип хромосомной мутации является наиболее распространенным среди эуплоидий и характеризуется тем, что в организме содержится три или более полных набора хромосом.
Организм (обычно диплоид = 2n) считается полиплоидным, если он приобретает один или несколько полных наборов хромосом. В отличие от точечных мутаций, хромосомных инверсий и дупликаций, этот процесс является крупномасштабным, то есть он происходит на полных наборах хромосом.
Вместо гаплоида (n) или диплоида (2n) полиплоидный организм может быть тетраплоидом (4n), октоплоидом (8n) или выше. Этот процесс мутации довольно часто встречается у растений и редко встречается у животных. Этот механизм может увеличить генетическую изменчивость сидячих организмов, которые не могут перемещаться.
Полиплоидия имеет большое значение с точки зрения эволюции в определенных биологических группах, где она представляет собой частый механизм для генерации новых видов, поскольку хромосомная нагрузка является наследственным заболеванием.
Когда возникает полиплоидия?
Нарушения числа хромосом могут возникать как в природе, так и в лабораторно установленных популяциях. Их также можно индуцировать мутагенными агентами, такими как колхицин. Несмотря на невероятную точность мейоза, хромосомные аберрации действительно происходят и встречаются чаще, чем можно было бы подумать.
Полиплоидия возникает в результате некоторых изменений, которые могут происходить во время мейоза, либо в первом мейотическом делении, либо во время профазы, когда гомологичные хромосомы организованы парами, образуя тетрады, и нерасхождение последних происходит во время анафаза I.
Появление новых видов
Полиплоидия важна, так как это отправная точка для возникновения новых видов. Этот феномен является важным источником генетической изменчивости, поскольку он дает начало сотням или тысячам дублирующих локусов, которые остаются свободными для получения новых функций.
В растениях это особенно важно и довольно широко распространено. По оценкам, более 50% цветковых растений произошли от полиплоидии.
В большинстве случаев полиплоиды физиологически отличаются от исходных видов и благодаря этому могут колонизировать среду с новыми характеристиками. Многие важные виды в сельском хозяйстве (включая пшеницу) представляют собой полиплоиды гибридного происхождения.
Типы полиплоидии
Полиплоидии можно классифицировать по количеству наборов или полных наборов хромосом, присутствующих в ядре клетки.
С другой стороны, полиплоидии также можно классифицировать по происхождению хромосомных данных. В этом порядке представлений организм может быть автополиплоидом или аллополиплоидом.
Автополиплоид содержит несколько наборов гомологичных хромосом, полученных от одного и того же человека или от человека, принадлежащего к одному виду. В этом случае полиплоиды образуются путем объединения невосстановленных гамет генетически совместимых организмов, занесенных в каталог как один и тот же вид.
Аллополиплоид - это организм, который содержит негомологичные наборы хромосом из-за гибридизации между разными видами. В этом случае полиплоидия возникает после гибридизации между двумя родственными видами.
Полиплоидия у животных
Полиплоидия у животных встречается редко. Наиболее распространенная гипотеза, объясняющая низкую встречаемость полиплоидных видов у высших животных, заключается в том, что их сложные механизмы определения пола зависят от очень тонкого баланса числа половых хромосом и аутосом.
Эта идея сохранилась, несмотря на накопление доказательств существования полиплоидов животных. Обычно это наблюдается у низших групп животных, таких как черви и широкий спектр плоских червей, у которых обычно есть гонады мужских и женских особей, что способствует самооплодотворению.
Виды с последним состоянием называются самосовместимыми гермафродитами. С другой стороны, это также может происходить в других группах, самки которых могут давать потомство без оплодотворения, посредством процесса, называемого партеногенезом (который не подразумевает нормальный мейотический половой цикл).
Во время партеногенеза потомство производится в основном путем митотического деления родительских клеток. Сюда входят многие виды беспозвоночных, такие как жуки, равноногие моли, бабочки, креветки, различные группы паукообразных и некоторые виды рыб, амфибий и рептилий.
В отличие от растений, видообразование посредством полиплоидии - исключительное явление для животных.
Примеры на животных
Полиплоидия у человека
Полиплоидия редко встречается у позвоночных и считается неуместной для диверсификации групп, таких как млекопитающие (в отличие от растений), из-за сбоев в системе определения пола и в механизме компенсации дозы.
По оценкам, пять человек из 1000 рождаются с серьезными генетическими дефектами, связанными с хромосомными аномалиями. Еще больше эмбрионов с хромосомными дефектами выкидывают самопроизвольно, а многие еще не доживают до рождения.
У человека хромосомные полиплоидии считаются летальными. Однако в соматических клетках, таких как гепатоциты, около 50% из них обычно полиплоидные (тетраплоидные или октаплоидные).
Полиплоидии, наиболее часто обнаруживаемые у нашего вида, - это полные триплоидии и тетраплоидии, а также диплоидные / триплоидные (2n / 3n) и диплоидные / тетраплоидные (2n / 4n) миксоплоиды.
В последнем случае популяция нормальных диплоидных клеток (2n) сосуществует с другой популяцией, которая имеет 3 или более гаплоидных кратных хромосом, например: триплоид (3n) или тетраплоид (4n).
Триплоидии и тетраплодии у человека не жизнеспособны в долгосрочной перспективе. В большинстве случаев сообщалось о смерти при рождении или даже в течение нескольких дней после рождения, в диапазоне от менее одного месяца до максимум 26 месяцев.
Полиплоидия у растений
Существование более чем одного генома в одном ядре сыграло важную роль в происхождении и эволюции растений, будучи, возможно, наиболее важным цитогенетическим изменением в видообразовании и эволюции растений. Растения были воротами к знаниям о клетках с более чем двумя наборами хромосом на клетку.
С самого начала подсчета хромосом было замечено, что большое количество диких и культурных растений (включая некоторые из наиболее важных) полиплоидны. Почти половина известных видов покрытосеменных (цветковых) являются полиплоидными, как и большинство папоротников (95%) и самые разнообразные мхи.
В пределах растений, возможно, аллополиплоиды (более распространенные в природе) играли фундаментальную роль в видообразовании и адаптивном излучении многих групп.
Улучшение садоводства
У растений полиплоидия может возникать в результате нескольких различных явлений, возможно, наиболее частыми из которых являются ошибки в процессе мейоза, приводящие к образованию диплоидных гамет.
Более 40% культурных растений являются полиплоидными, включая люцерну, хлопок, картофель, кофе, клубнику, пшеницу, среди прочего, без связи между одомашниванием и полиплоидией растений.
Поскольку колхицин применялся как агент, вызывающий полиплоидию, он использовался в сельскохозяйственных культурах в основном по трем причинам:
-Для полиплоидизации гибридов и восстановления фертильности таким образом, чтобы некоторые виды были переработаны или синтезированы.
-И, наконец, как способ передачи генов между видами с разной степенью плоидности или внутри одного и того же вида.
Примеры в растениях
Пшеница в культурных растениях играет исключительно важную роль. Существует 14 видов пшеницы, которые эволюционировали путем аллополиплоидии, и они образуют три группы: одну из 14, другую из 28 и последнюю из 42 хромосом. В первую группу входят самые старые виды рода. Т. monococcum Y T. boeoticum.
Вторая группа состоит из 7 видов и, по-видимому, происходит от гибридизации T. boeoticum с видом дикорастущих трав другого рода, называемого Эгилопс. Скрещивание дает сильный стерильный гибрид, который в результате дупликации хромосом может привести к фертильному аллотетраплоиду.
Третья группа из 42 хромосом - это хлебная пшеница, которая, вероятно, возникла в результате гибридизации тертраплоидного вида с другим видом Эгилопс с последующим удвоением хромосомного набора.
Прочитав эту статью, вы узнаете, что такое полиплоидия. Мы рассмотрим, какую роль она играет. Вы также узнаете, какие бывают виды полиплоидии.
Образование полиплоидов
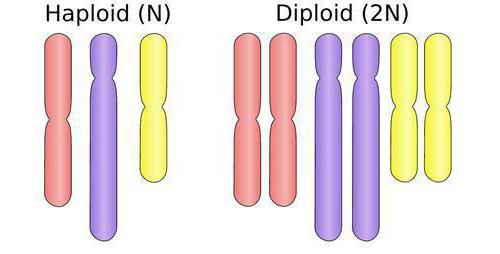
Свойства полиплоидов
У этих растений изменчивость часто бывает значительно уже, чем у родственных диплоидов, поскольку каждый ген представлен у них по меньшей мере в удвоенном числе. При расщеплении в потомстве гомозиготные по какому-нибудь рецессивному гену особи составят только 1/16 вместо 1/4 у диплоидов. (В обоих случаях принимается, что частота рецессивных аллелей равна 0,50.) Полиплоидам свойственно самоопыление, еще более снижающее их изменчивость, несмотря на то что родственные им диплоиды преимущественно опыляются перекрестно.
Где встречаются полиплоиды
Итак, мы ответили на вопрос, что такое полиплоидия. А где же встречаются такие растения?
Одни полиплоиды лучше приспособлены к сухим местам или более низким температурам, чем исходные диплоидные формы, в то время как другие лучше приспособлены к особым типам почв. Благодаря этому они могут заселять места с экстремальными условиями существования, в которых их диплоидные предки, скорее всего, погибли бы. С небольшой частотой они встречаются во многих естественных популяциях. Они легче, чем соответствующие им диплоиды, вступают в неродственные скрещивания. При этом сразу же могут получаться плодовитые гибриды. Реже полиплоиды гибридного происхождения образуются путем удвоения числа хромосом у стерильных диплоидных гибридов. Это один из путей восстановления плодовистости.
Первый документированный случай полиплоидии
Именно таким, менее обычным, путем образовались полиплоидные гибриды между редькой и капустой. Это был первый хорошо задокументированный случай полиплоидии. Оба рода принадлежат к семейству крестоцветных и находятся в близком родстве. В соматических клетках и того и другого вида находится 18 хромосом, и в первой метафазе мейоза всегда обнаруживается 9 пар хромосом. С некоторым трудом был получен гибрид между этими растениями. В мейозе он имел 18 непарных хромосом (9 от редьки и 9 от капусты) и был совершенно бесплоден. Среди этих гибридных растений спонтанно образовался полиплоид, у которого в соматических клетках было 36 хромосом и в процессе мейоза регулярно образовывались 18 пар. Иными словами, полиплоидный гибрид имел все 18 хромосом как редьки, так и капусты, и они функционировали нормально. Этот гибрид был довольно плодовитым.
Полиплоиды-сорняки
Некоторые полиплоиды возникали как сорняки в местах, связанных с деятельностью человека, и иногда они достигали удивительного процветания. Один из хорошо известных примеров — обитатели соленых болот из рода Spartina. Один из видов, S. maritima (на фото ниже), встречается на болотах вдоль берегов Европы и Африки. Другой вид, S. alterniflora, был завезен в Великобританию с востока Северной Америки около 1800 г. и впоследствии широко распространился, образовав крупные локальные колонии.

Пшеница
Одной из важнейших полиплоидных групп растений можно считать род Triticum пшеницы (на фото далее). Самая распространенная в мире хлебная культура — мягкая пшеница (Т. aestivum) — имеет 2n = 42. Мягкая пшеница возникла как минимум 8000 лет назад, вероятно, в Центральной Европе, в результате естественной гибридизации возделываемой пшеницы, имеющей 2n = 28, с диким злаком того же рода, имеющим 2n = 14. Дикий злак, вероятно, рос как сорняк среди посевов пшеницы. Гибридизация, давшая начало мягкой пшенице, могла произойти между полиплоидами, появлявшимися время от времени в популяциях обоих родительских видов.
Triticosecale

Многообразие полиплоидов
В природе они отбираются под влиянием внешних условий, а не благодаря деятельности человека. Их возникновение — один из важнейших эволюционных механизмов. В наше время множество полиплоидов представлено в мировой флоре (более половины всех видов растений). Среди них многие из наиболее важных сельскохозяйственных культур — не только пшеница, но и хлопчатник, сахарный тростник, банан, картофель и подсолнечник. К этому перечню можно добавить большинство красивых садовых цветов — хризантемы, анютины глазки, георгины.
Теперь вы знаете, что такое полиплоидия. Ее роль в сельском хозяйстве, как вы видите, очень велика.
Читайте также: