
Образование макроспор образование гаметофита двойное оплодотворение
Обновлено: 04.07.2024
Микроспорогенез или образование мужского гаметофита. Взрослая тычинка состоит из пыльника, связника и тычиночной нити, которая одета эпидермой. Тычинка образуется из бугорка конуса нарастания цветоносного побега, который сначала однороден, а затем дифференцируется. Постепенно в двух участках каждого пыльцевого мешка начинается заложение гнезд пыльника; выделяется археспориальная клетка, а она делится, образуя париетальную и спорогеннуюклетки.
Париетальные клетки дают субэпидермальный фиброзный слой и выстилающий – тапетум, обеспечивающий спорогенную ткань питательными веществами. Цитоплазма густая, вязкая, клетки крупные.
Спорогенные клетки, делясь мейозом, дают клетки микроспор, в дальнейшем дифференцирующихся в пыльцу.
Ядро микроспоры делится на вегетативное и генеративное ядра.
Вегетативное - крупное, содержащее жирное масло и крахмал.
Генеративное - мельче, с хроматином.
Генеративное может делиться еще в пыльнике, образуя два спермия - (сложноцветные, злаки, маревые, гвоздичные), или же в пыльцевой трубке (орхидные, норичниковые).
Следовательно, зрелые пыльцевые зерна могут быть двух- и трехъядерные, покрыты спородермой, состоящей из экзины и интины Экзина - утолщенная слоистая оболочка, с выростами в виде шипиков, бугорков и т.д. Состоит из вещества - полленина, клетчатки, пропитанной кутиноподобными веществами, маслом с каротиноидами.
Интинасостоит из пектиновых веществ.
Экзина имеет поры. Форма и характер поверхности экзины - диагностический признак.
Вскрывание пыльников обусловлено, в основном, сокращением клеток эпидермы при подсыхании.
Мегаспорогенез или формирование женского гаметофита. Семяпочки развиваются на внутренних стенках завязи и являются мегаспорангиями, заключенными в интегументы. Закладываются в виде бугорков на плаценте (место прикрепления семяпочки). Главная масса семяпочки - эмбриональные клетки, образующиеся из бугорков, формирующие нуцеллус (питательная и защитная ткань зародышевого мешка). Нуцеллус покрыт интегументами, оставляющими узкий канал в виде пыльцевхода (микропиле). Часть семяпочки, противоположная пыльцевходу – халаза.
Количество семяпочек в завязи неодинаково (слива, вишня - 1-2; злаки - 1; мак - много), соответственно этому неодинаково и число семян.
Форма зародышевого мешка разнообразна: овальный ,прямой, изогнутый и т.п. В цитоплазме зародышевого мешка имеются лейкопласты, хондриосомы, крахмальные зерна, капли масла и т.д.
Оплодотворение.После опыления начинается прорастание пыльцы. Липкая и неровная поверхность рыльца способствует удержанию большого числа пыльцевых зерен. Кроме того, если партнеры совместимы, рыльце выделяет специальный энзим, стимулирующий прорастание пыльцы. Прорастает несколько пыльцевых зерен. От индивидуальной скорости роста зависит успех той или иной пыльцы. В пыльцевую трубку переходят два спермия и вегетативное ядро, которое располагается у растущего конца трубки. Трубка входит в семяпочку через микропиле, конец ее лопается и содержимое изливается в полость зародышевого мешка. Один спермий сливается с гаплоидной яйцеклеткой, другой с диплоидной вторичной клеткой.
Обычно в зародышевый мешок проникает одна пыльцевая трубка, если семяпочек много, то несколько выполняют свое назначение.
Из диплоидной зиготы развивается зародыш семени, из триплоидной - эндосперм. Описанный универсальный для покрытосеменных процесс - двойное оплодотворение, открыт Сергеем Гавриловичем Навашиным в 1898 году.
От голосеменных отличия:
1) гибридный, триплоидный эндосперм;
2) два (а не один) спермий.
3) время от опыления до оплодотворения - 1-2 дня.
Смена ядерных фаз и чередование поколений. Женский гаметофит - зародышевый мешок.
Мужской гаметофит - пыльца.
Обыкновенный тип двойного оплодотворения - амфимиксис. Но у некоторых растений зародыш развивается из неоплодотворенных элементов. Отклонения от нормального хода оплодотворения - апомиксис. Различают:
1. Партеногенез –развитие зародыша из неоплодотворенной яйцеклетки;
2. Апогамия - развитие зародыша из антиподы или синергиды;
3. Апоспория - зародыш развивается из клетки нуцеллуса или интегумента.
После оплодотворения начинается деление оплодотворенных элементов внутри зародышевого мешка. Триплоидная зигота образует эндосперм, раньше зародыша. Зародыш развивается из зиготы. Может быть и перисперм - из нуцеллуса (вне зародышевого мешка). Интегументы становятся кожурой семени.
Зародыш, эндосперм, перисперм и кожура образуют семя .
Различают 4 типа семян.
1. Зародыш занимает всю полость зародышевого мешка, вытесняя весь эндосперм и перисперм. Питательные вещества - в семядолях (бобовые, тыквенные, розоцветные, сложноцветные).
2. Зародыш занимает меньшую часть семени, большая принадлежит эндосперму. Перисперма не образуется (злаки, пасленовые, зонтичные).
3. Есть только перисперм. Эндосперм полностью потреблен зародышем в процессе формирования (гвоздичные, маревые).
4.Сохраняется эндо- и перисперм - редкий тип семян (черный перец, лотос).
При образовании зародыша зигота делится на две клетки, образуя
1) подвесок или суспензор,
2) собственно зародыш.
Подвесок погружает зародыш в эндосперм, нередко выполняет функции присоски.
Зародыш состоит из зародышевого корешка, стебелька, к верхней части которого прикрепляются семядоли - первые листья растения. Стебелек ниже семядолей - гипокотиль, выше - эпикотиль; верхушка стебля - почечка.
© 2014-2022 — Студопедия.Нет — Информационный студенческий ресурс. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав (0.004)
Двойное оплодотворение это сложный оплодотворение механизм цветения растений (покрытосеменные). Этот процесс предполагает присоединение самки гаметофит (мегагаметофит, также называемый зародышевый мешок) с двумя самцами гаметы (сперма). Это начинается, когда пыльца придерживается стигмы карпель, женская репродуктивная структура цветка. Затем пыльцевое зерно впитывает влагу и начинает прорастать, образуя пыльцевая трубка что простирается вниз к яичник через стиль. Затем кончик пыльцевой трубки входит в яичник и проходит через микропиле отверстие в семяпочке. Пыльцевая трубка продолжает выпуск двух сперматозоидов мегагаметофита.
В неоплодотворенной семяпочке 8 клеток, расположенных в виде 3 + 2 + 3 (сверху вниз), то есть 3 антиподальных клетки, 2 полярных центральных клетки, 2 синергид и 1 яйцеклетка. Один сперматозоид оплодотворяет яйцеклетку, а другой сперматозоид соединяется с двумя полярные ядра из большого центральная ячейка мегагаметофита. Гаплоидная сперма и гаплоидное яйцо объединяются, чтобы сформировать диплоид зигота, процесс, называемый сингамия, в то время как другая сперма и два гаплоидный полярные ядра крупной центральной клетки мегагаметофита образуют триплоид ядро (тройной сплав). Некоторые растения могут образовывать полиплоид ядра. Большая клетка гаметофита затем разовьется в эндосперм, богатая питательными веществами ткань, обеспечивающая питание развивающегося эмбриона. Завязь, окружающая семяпочки, превращается в плод, который защищает семена и может их рассеивать. [1]
Два материнских ядра центральной клетки (полярные ядра), которые вносят вклад в эндосперм, возникают в результате митоза из одного и того же единственного ядра. мейотический продукт, из которого выросло яйцо. Материнский вклад в генетическую конституцию триплоидного эндосперма вдвое больше, чем вклад эмбриона.
В исследовании, проведенном в 2008 г. Arabidopsis thaliana, миграция мужских ядер внутри женской гаметы в слиянии с женскими ядрами была впервые задокументирована с использованием in vivo визуализация. Некоторые из генов, участвующих в процессе миграции и слияния, также были определены. [2]
Свидетельства двойного оплодотворения в Gnetales, которые являются нецветущими семенными растениями. [3]
Содержание
Краткая история
Двойное оплодотворение было открыто более века назад Сергей Наващин и Гриньяр в Киев, [4] Российская империя, и Леон Гиньяр в Франция. Каждый сделал открытие независимо от другого. [5] Лилиум мартагон и Фритиллярия Tenella были использованы в первых наблюдениях двойного оплодотворения, которые были сделаны с использованием классической оптический микроскоп. Из-за ограничений светового микроскопа оставалось много безответных вопросов относительно процесса двойного оплодотворения. Однако с развитием электронный микроскоп, на многие вопросы были даны ответы. В частности, наблюдения, сделанные группой У. Дженсена, показали, что мужские гаметы не имеют никаких клеточные стенки и что плазматическая мембрана гамет находится близко к плазматической мембране клетки, которая окружает их внутри пыльцевого зерна. [6]
Двойное оплодотворение голосеменных растений
Двойное оплодотворение in vitro
Связанные структуры и функции
Мегагаметофит
Женский гаметофит, мегагаметофит, который участвует в двойном оплодотворении в покрытосеменные который является гаплоидным, называется зародышевым мешком. Это развивается в яйцеклетка, окруженная яичником у основания карпель. Мегагаметофит окружают (один или) два кожные покровы, которые образуют отверстие, называемое микропиле. Мегагаметофит, который обычно гаплоидный, происходит от (обычно диплоид) мегаспора материнская ячейка, также называемая мегаспороцит. Следующая последовательность событий варьируется в зависимости от конкретного вида, но у большинства видов происходят следующие события. Мегаспороцит подвергается мейотическому делению клеток, образуя четыре гаплоидных мегаспоры. Только одна из четырех образующихся мегаспор выживает. Эта мегаспора проходит три раунда митотического деления, в результате чего образуется семь клеток с восемью гаплоидными ядрами (центральная клетка имеет два ядра, называемые полярными ядрами). Нижний конец эмбрионального мешка состоит из гаплоидной яйцеклетки, расположенной в середине двух других гаплоидных клеток, называемых синергиды. Синергиды действуют в привлечении и направлении пыльцевой трубки к мегагаметофиту через микропиле. На верхнем конце мегагаметофита находятся три антиподальные клетки.
Микрогаметофит
Мужские гаметофиты или микрогаметофиты, участвующие в двойном оплодотворении, содержатся внутри пыльца зерна. Они развиваются внутри микроспорангии или мешочки с пыльцой, из пыльников тычинок. Каждый микроспорангий содержит диплоид микроспора материнские клетки или микроспороциты. Каждый микроспороцит подвергается мейозу, образуя четыре гаплоидных микроспоры, каждая из которых в конечном итоге может развиться в пыльцевое зерно. Микроспора подвергается митоз и цитокинез чтобы произвести две отдельные клетки, генеративную клетку и трубочную клетку. Эти две клетки помимо стенки спор составляют незрелое пыльцевое зерно. По мере созревания мужского гаметофита генеративная клетка переходит в трубочную клетку, а генеративная клетка подвергается митозу, производя две сперматозоиды. Как только пыльцевые зерна созреют, пыльники взломать, выпуская пыльцу. Пыльца попадает в пестик другого цветка, ветром или животными-опылителями, и отложился на рыльце. По мере прорастания пыльцевых зерен трубчатая ячейка производит пыльцевую трубку, которая удлиняется и простирается вниз по длине плодолистика до яичника, где его сперматозоиды высвобождаются в мегагаметофит. Отсюда происходит двойное оплодотворение. [18]
рганом полового размножения покрытосеменных растений является цветок. Цветок – видоизмененный, укороченный, неразветвленный побег, предназначенный для образования спор и гамет и полового процесса, завершающегося образованием семян и плода.
Растение, на котором образуются споры, называют спорофит . У сосудистых растений (хвощи, плауны, папоротники, семенные растения) спорофит – листостебельное диплоидное растение. Цветковые – разноспоровые растения образующие споры, отличающиеся по величине и физиологическим особенностям, образуются споры в результате мейоза.
Микроспоры – более мелкие споры, формирующиеся в тычинках, из них образуются пыльцевые зерна – мужские гаметофиты , образующие мужские гаметы. В семязачатке, в нуцеллусе (мегаспорангии) в результате мейоза образуется мегаспора – более крупная спора, из нее образуется женский гаметофит, названный зародышевым мешком . После опыления и двойного оплодотворения образуется плод и семена.
И образование спор, и слияние гамет происходит в цветке.
Строение цветка. У цветка различают цветоножку, цветоложе, околоцветник, тычинки и пестики. У некоторых цветков отдельные части могут отсутствовать.
1 — цветоножка; 2 — цветоложе; 3 — чашелистики; 4 лепесток; 5 — пестик; 6 — тычинка
ветки большинства видов растений имеют и тычинки, и пестики. Такие цветки называют обоеполыми (вишня, горох). Цветки, которые имеют только пестики, называют пестичными (женскими). Цветки, которые имеют только тычинки, называют тычиночными (мужскими). В зависимости от распределения однополых цветков на растениях различают: однодомные растения – растения, у которых на одних и тех же экземплярах располагаются и женские, и мужские цветки (огурец, кукуруза, дуб); двудомные растения – растения, у которых на одних экземплярах располагаются женские, а на других – мужские цветки (крапива двудомная, конопля, облепиха); многодомные растения – растения, у которых на одних и тех же экземплярах встречаются как обоеполые, так и однополые цветки в различных количественных соотношениях (гречиха, некоторые виды ясеня, клена).
Цветоножка – междоузлие под цветком. Цветки, лишенные цветоножки, называются сидячими (цветки в соцветии корзинка у подсолнечника, астры, одуванчика). Цветоложе – укороченная стеблевая часть цветка. На ней располагаются все остальные части цветка.
Околоцветник – стерильная часть цветка, его покров. Околоцветник может быть простым (не дифференцированным на чашечку и венчик, образованным совокупностью одинаковых листочков) и двойным (дифференцированным на чашечку и венчик). Простой околоцветник может быть венчиковидным (образованным ярко окрашенными листочками), или чашечковидным (образованным зелеными листочками). Цветки, лишенные околоцветника (ива, тополь), называются голыми.
Чашечка – наружная часть двойного околоцветника, представляет собой совокупность чашелистиков – видоизмененных прицветных листьев Обычно чашелистики имеют небольшие размеры и зеленую окраску. Венчик – внутренняя, обычно окрашенная часть двойного околоцветника. Представляет собой совокупность лепестков, часто имеющих яркую окраску.
Андроцей (дом для мужчин) – совокупность тычинок одного цветка. У большинства растений тычинка состоит из тычиночной нити и пыльника. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда ( микроспорангия ), в которых происходит образование микроспор, а впоследствии пылинок.
Микроспорогенез – процесс образования микроспор в микроспорангиях (гнездах пыльника). Микроспоры формируются из материнских клеток – микроспороцитов , имеющих диплоидный набор хромосом. В результате мейоза каждая материнская клетка образует четыре гаплоидных микроспоры. Микроспоры быстро обособляются друг от друга.
Рис. . Образование мужского гаметофита
1 – микроспороцит (2n); 2 – микроспоры (n); 3 – вегетативная клетка; 4 – генеративная клетка; 5 – экзина; 6 – интина; 7 – 2 спермия.
икрогаметогенез – процесс образования мужских половых клеток (спермиев), происходит в пыльцевом зерне, которое является мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита происходит также в гнездах пыльников тычинок и сводится к одному митотическому делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины (наружной, толстой). Каждой пыльцевое зерно содержит две гаплоидные клетки: вегетативную и генеративную . Из генеративной (спермагенной) далее образуются два спермия . Из вегетативной (сифоногенной) впоследствии образуется пыльцевая трубка.
Гинецей (дом для женщин) – совокупность пестиков одного цветка. Обычно в пестике выделяют три части: завязь, столбик и рыльце. Завязь – замкнутая, нижняя, полая часть пестика, несущая и защищающая семязачатки. В завязи может располагаться от одного (пшеница, вишня) до нескольких тысяч (мак) семязачатков. Стенки завязи выполняет функцию защиты семязачатков от неблагоприятных факторов среды (высыхание, колебание температур, поедание насекомыми и т.д.), внутри завязи (в семязачатках) происходит мегаспорогенез и мегагаметогенез, они принимают участие в образовании околоплодника.
Семязачаток состоит из нуцеллуса (ядра) – центральной части, являющейся мегаспорангием, двух покровов – интегументов, которые при смыкании образуют узкий канал – микропиле, или пыльцевход, через который пыльцевая трубка проникает к зародышевому мешку. С помощью семяножки семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где сливаются нуцеллус и интегументы, называют халазой.
1 – интегументы; 2 – микропиле; 3 – плацента; 4 – семяножка; 5 – нуцеллус; 6 – синергиды; 7 – яйцеклетки; 8 – центральная клетка; 9 – антиподы; 10 – халаза.
семязачатке происходит мегаспорогенез, мегагаметогенез и процесс оплодотворения. После оплодотворения (реже без него) из семязачатка формируется семя.
Мегаспорогенез – процесс формирование мегаспор. Он происходит в нуцеллусе семязачатка. После заложения семязачатка и формирования нуцеллуса в области микропиле начинает разрастаться одна клетка – мегаспороцит , или материнская клетка мегаспор . Материнская клетка мегаспор имеет диплоидный набор хромосом. У большинства покрытосеменных из нее путем мейоза образуются четыре гаплоидные мегаспоры. Из четырех мегаспор лишь одна дает начало женскому гаметофиту – зародышевому мешку. Остальные мегаспоры отмирают.
Рис. . Образование женского гаметофита:
1 – мегаспороцит (2n); 2 – четыре мегаспоры; 3 – разрастание одной из мегаспор и отмирание трех остальных; 4 – антиподы; 5 – образование центральной клетки (2n); 6 – яйцеклетка; 7 – синергиды.
егагаметогенез – процесс формирования женских половых клеток, происходит в зародышевом мешке. Формирование женского гаметофита начинается с разрастания мегаспоры, которая далее три раза делится митозом. В результате этого образуются восемь клеток, которые располагаются следующим образом: три – на одном полюсе зародышевого мешка (микропилярном), три – на другом (халазальном), две – в центре. Две оставшиеся в центре клетки сливаются, образуя диплоидную центральную клетку зародышевого мешка. Одна из трех клеток, расположенных на микропилярном полюсе, отличается большими размерами и является яйцеклеткой . Две рядом расположенные клетки являются вспомогательными и называются синергидами . Группа из трех клеток, находящихся на противоположном, халазальном полюсе, называется антиподами . Таким образом, сформированный женский гаметофит включает шесть гаплоидных клеток (яйцеклетка, две клетки-синергиды, три клетки-антиподы) и одну диплоидную клетку.
Оплодотворение . Образование семян и плодов. Процессу оплодотворения предшествует опыление – перенос пыльцы от пыльцевых мешков тычинок к рыльцам пестиков. Попав на рыльце пестика, под воздействием веществ, выделяемых пестиком, пыльца начинает прорастать: образуется пыльцевая трубка, внедряющаяся в ткань рыльца. Кончик пыльцевой трубки выделяет вещества, размягчающие ткань рыльца и столбика. В процессе формирования пыльцевой трубки принимает участие сифоногенная клетка. По мере роста пыльцевой трубки в нее переходит спермагенная клетка, которая делится митозом с образованием двух спермиев (у некоторых растений спермагенная клетка дает начало двум спермиям еще до прорастания пыльцы). Пыльцевая трубка продвигается по столбику пестика и врастает в зародышевый мешок, как правило, через микропиле. После проникновения в зародышевый мешок кончик пыльцевой трубки разрывается, и спермии попадают внутрь. Один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу. Второй спермий сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку, из которой далее формируется эндосперм (питательная ткань) семени, обеспечивающий питание зародыша. Синергиды и антиподы дегенерируют. В целом выше описанный процесс получил название двойного оплодотворения. Двойное оплодотворение у цветковых растений было открыто в 1898 году русским ботаником С.Г.Навашиным.
После двойного оплодотворения из яйцеклетки формируется зародыш семени, из центрального ядра зародышевого мешка – триплоидный эндосперм, из интегументов – семенная кожура, из всего семязачатка – семя, а из стенок завязи – околоплодник. В целом из завязи пестика формируется плод с семенами .
Ключевые термины и понятия
1. Спорофит. 2. Разноспоровые растения. 3. Пестичные и тычиночные цветки. 4. Однодомные, двудомные и многодомные растения. 5. Андроцей. 6. Гинецей. 7. Микроспорогенез. 8. Микрогаметогенез. 9. Мегаспорогенез. 10. Мегагаметогенез. 11. Мужской гаметофит. 12. Женский гаметофит. 13. С.Г.Навашин.

Видеоурок способствует формированию представлений о способах опыления цветковых растений, выявляет приспособления к различным способам опыления. Основная цель урока – рассмотреть особенности двойного оплодотворения и его преимущества. В ходе видеоурока подробно рассказывается о процессах микроспорогенеза и макроспорогенеза, их сущности.

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Двойное оплодотворение покрытосеменных растений"
Размножение – это одно из обязательных свойств любого живого организма. Оно заключается в увеличении числа особей.
Различают бесполое и половое размножение растений.
Бесполое размножение подразделяют на спорообразование и вегетативное.
Половое размножение происходит при помощи особых половых клеток –гамет.
При бесполом размножении быстро увеличивается численность вида, все потомки имеют абсолютно такой же генотип, что и родительская особь. А также не происходит увеличения генетического разнообразия, которое может оказаться очень полезным при изменении условий существования вида.
По этой причине большинство живых организмов на Земле размножаются половым путём.
Сущность полового размножения заключается в слиянии генетической информации родителей, благодаря чему генетическое разнообразие в потомстве увеличивается.
У покрытосеменных растений половое размножение связано с цветком.
Важные части цветка – это пестик и тычинка. С их участием происходят сложные процессы полового размножения –опыление и оплодотворение.
Но сперва в будущем цветке начинают образовываться половые клетки.
В жизненном цикле цветковых растений наблюдается смена поколений.
У растений выделяют диплоидное поколение – бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Гаметы образуются в результате митоза, а споры – в результате мейоза. И гаметы, и споры образуются в цветке, поэтому цветок является органом и бесполого, и полового размножения.
Пыльцевое зерно является спорой (микроспорой), а не мужской гаметой, так как в нем самом развиваются мужские гаметы.
У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток.
Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае из 7 клеток (содержит 8 – либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра).
Посмотрим, как образуется мужской гаметофит.
Итак, Микроспорогенез
В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делений становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, поверхность её либо гладкая, либо шероховатая, приспособленная для переноса пыльцы и прилипания её к рыльцу пестика.
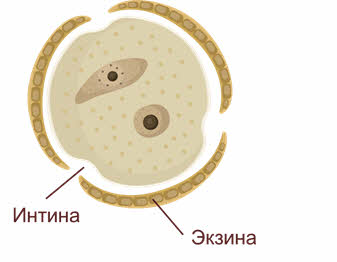
Этим заканчивается микроспорогенез, вслед за образованием одноядерной микроспоры начинается микрогаметогенез.
Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и её ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться ещё в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, называются спермиоклетками, или спермиями.
Таким образом, из одной споры (микроспоры) с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: два из них – спермии и одно – вегетативное. При образовании пыльцевой трубки это вегетативное ядро переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены российским и советским цитологом и эмбриологом растений Сергеем Гавриловичем Навашиным в 1910 г. на лилейных растениях.
После образования гамет пыльник созревает, и пыльца высыпается. Она несёт только генетическую информацию.
Посмотрим, как происходит мегаспорогенез и мегагаметогенез цветковых.
У покрытосеменных растений женский гаметофит – это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез.
В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растёт, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют, судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем её ядро претерпевает ряд делений. При этом сама клетка не делится, а делится только ядро.
У разных систематических групп растений число делений ядра мегаспоры может варьировать от одного до трёх. У большинства растений (70 % видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других – в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
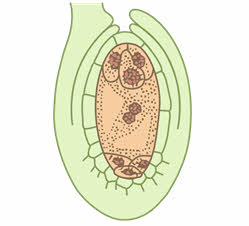
В дальнейшем от каждой из двух полярных четвёрки ядер к центру отходит по одному ядру, которые сливаются, образуя вторичное (центральное) ядро зародышевого мешка. Затем цитоплазма обособляется вокруг ядер гаметофита, который из ядерной стадии развития переходит в клеточную. Три ядра, оставшиеся вблизи халазального полюса, преобразуются в три клетки (антиподы), которые питают гаметофит. Три ядра вблизи микропиле отделяются клеточными перегородками, образуя отдельные клетки: крупную центральную яйцеклетку и две боковые клетки синергиды. Вся цитоплазма, расположенная между антиподами с одной стороны и клетками яйцевого комплекса с другой (яйцеклетка и две синергиды), называется центральной клеткой. В ней находится диплоидное вторичное (центральное) ядро.
На этом этапе женский гаметофит уже полностью сформирован и состоит из шести гаплоидных клеток (одной яйцеклетки, двух синергид и трёх антипод) и одной диплоидной (центральной). Его строение внешне напоминает мешочек, поэтому женский гаметофит покрытосеменных называется зародышевым мешком.
Пылинка попадает на рыльце пестика, и происходит опыление.
Опыление – это перенос пыльцевых зёрен на рыльце пестика, у голосеменных пыльцевые зерна при опылении попадают непосредственно на семязачаток.
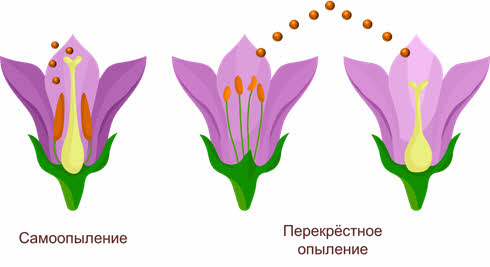
Имеется два основных типа опыления: самоопыление (автогамия) (когда растение опыляется собственной пыльцой) и перекрёстное опыление (аллогамия).
При самоопылении исключён обмен генетической информацией, поскольку пыльцевые зерна попадают на рыльце пестика либо с одной из тычинок этого же цветка, либо с другого цветка, расположенного на том же растении. Это приводит к появлению чистых линий гомозиготных популяций в пределах одного вида, неспособных обмениваться мутировавшими генами, поэтому процессы видообразования в этих популяциях идут самостоятельно.
Перекрёстное опыление – это перенос пыльцы одного растения на рыльце другого. Этот тип опыления встречается более часто, чем самоопыление, между разными особями одного вида происходит обмен аллелями, что приводит к увеличению доли гетерозиготных организмов.
Безусловно, перекрёстное самоопыление имеет большие преимущества по сравнению с самоопылением, поскольку возникшие мутации свободно распространяются в пределах популяции.
Способы опыления у перекрёстноопыляемых цветковых растений весьма разнообразны. Их можно разделить на две группы. Первая: перенос пыльцы осуществляется главным образом насекомыми, а также некоторыми позвоночными (птицами и летучими мышами). Соответственно, различают энтомофилию, орнитофилию и зоофилию. Растения, опыляемые животными, обычно имеют яркоокрашенные крупные цветки. Мелкие цветки, как правило, собраны в соцветия, что зрительно их увеличивает. Для привлечения опылителей служит нектар или большое количество пыльцы, которую опылители охотно поедают.
Вторая группа: пыльца переносится абиотическими факторами – ветром и реже водой, в связи с чем различают анемофилию и гидрофилию.
После попадания пыльцы на рыльце начинается прорастание пыльцевого зерна.
Экзина мужского гаметофита прорывается в области борозды или поры прорастания, и начинает формироваться пыльцевая трубка, которая растёт, проникает в рыльце, через столбик движется по направлению к завязи, где находится семязачаток.
Трубка растёт на кончике. В растущую пыльцевую трубку из пыльцевого зерна перемещается ядро клетки-трубки, а также спермин.
Обычно развитие пыльцевой трубки происходит при уже сформированных семязачатках в завязи. Достигнув завязи, пыльцевая трубка через микропиле проникает внутрь одного из находящихся там семязачатков.
Там пыльцевая трубка направляется к яйцевому аппарату, проникает в одну из синергид и освобождает спермин. Один из спермиев сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами центральной клетки, образуя триплоидное ядро (с тройным набором хромосом). Зигота даёт начало зародышу, а из триплоидной центральной клетки образуется ткань эндосперма.
Таким образом оплодотворение, при котором одна мужская гамета сливается с яйцеклеткой, а вторая ― с вторичным ядром, называется двойным оплодотворением.
Механизм оплодотворения цветковых –двойное оплодотворение, открыто отечественным учёным Сергеем Гавриловичем Навашиным в 1898 году.
Таким образом, оплодотворённый семязачаток развивается в семя, из зиготы возникает зародыш, из триплоидной центральной клетки – эндосперм, а из внешней части семязачатка (интегументов) образуется семенная кожура, из стенок завязи цветка – стенки плода.
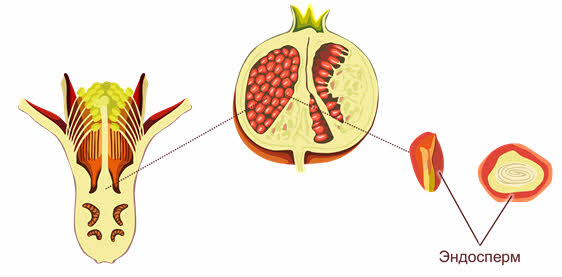
Эндосперм цветковых полностью отличается от первичного эндосперма голосеменных. У них первичный эндосперм представляет собой гаплоидную вегетативную ткань женского гаметофита, где накапливаются питательные вещества семени.
Эндосперм у покрытосеменных происходит из триплоидной центральной клетки, в образовании которой участвуют вторичное ядро женского гаметофита и ядро спермия.
У одних цветковых (например, злаков) эндосперм сильно разрастается и занимает большую часть семени, оттесняя на периферию маленький зародыш.
У других (к пример у бобовых) весь эндосперм поглощается зародышем и используется его семядолями, которые становятся самыми большими структурами зрелого семени.
Читайте также: