
Листорасположение у мхов всегда
Обновлено: 07.07.2024
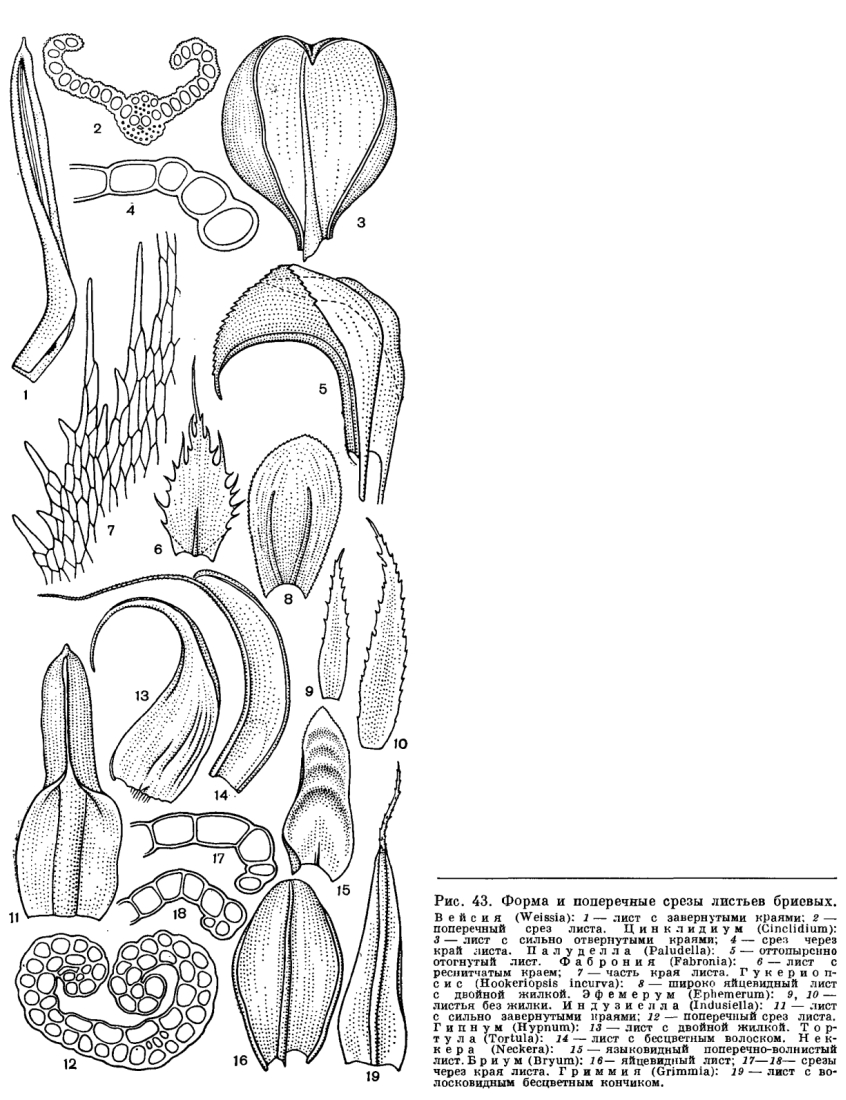
Листья у мхов сидячие, большей частью поперечно прикрепленные к стеблю, простые, цельные, по краю могут быть зубчатыми и только очень редко глубоко разделенными (рис. 43).
На стебле листья расположены всегда по спирали, образуя двурядное или многорядное листорасположение. Закладываются листья в восходящей последовательности. Рост их начинается с деления двусторонней верхушечной клетки, от которой отделяются сегменты с двух сторон. После прекращения верхушечного роста листья достигают конечного размера посредством вставочного роста, происходящего в их основании.
По расположению на стебле различают листья низовые, срединные (или собственно стеблевые) и покровные.
Покровпые листья окружают гаметангии и обычно отличаются от стеблевых размерами, формой, часто анатомическим строением, а иногда - окраской и консистенцией. Листья, окружающие архегонии, носят название перихециальных, а листья, окружающие антеридии,- перигониальных.
Низовые листья - сильно редуцированные, чешуевидные. Они развиваются в нижней надземной или в подземной части стебля у таких мхов, снабженных ризомом, как климациум (Climacium, табл. 8). Стеблевые листья расположены выше низовых, преимущественно в средней и верхпей частях стебля.
Листовая пластинка чаще однослойная, реже целиком или частично дву-, многослойная. Клетки пластинки обычно богаты хлоропластами и выполняют функцию фотосинтеза. Жилка, если она развита, обычно проходит посередине листа и состоит из толстостенных, вытянутых клеток. Кроме механической роли, она обеспечивает проведение пластических веществ и воды.
По форме листья очень разнообразны (рис. 43). Чрезвычайно различно строение краев листьев. Завернутость и отвернутость краев листа создает микроскопические полости, где и удерживается капиллярная вода. Завернутый край листа, свойственный мхам, обитающим в условиях периодической засухи, выполняет еще и защитную функцию, предохраняя нежные части листа от иссушения.
В листовой пластинке различают клетки двух основных типов: клетки паренхиматические - обычно округлые или угловатые до многоугольных (часто квадратные и шестиугольные), почти равной длины и ширипы, и прозенхиматические - узкие, вытянутые в длину с заостренными, заходящими друг за друга концами.
У мхов вся пластинка листа редко бывает построена из одинаковых клеток. Если верхняя часть листа состоит из паренхиматических мелких клеток, более толстостенных, а иногда мамиллозных или папиллозных, то клетки листового основания почти всегда несколько крупнее и резко отличаются от верхних по форме.
Большой полиморфизм клеток мхов обусловлен не только разнообразием их формы, но и характером их клеточных оболочек. Здесь их многообразие, кажется, не знает пределов.
Кроме разрастания боковых стенок клеток пластинки листа, могут разрастаться и наружные стенки, образуя мамиллозность и папиллозность (рис. 43). Папиллы (утолщения клеточной оболочки) встречаются наиболее часто у бриевых мхов. Они бывают чрезвычайно разнообразной формы, а иногда к тому же в месте их образования клеточная оболочка разрастается и выпячивается. На этом образовании и сидят папиллы. Папиллы сильно увеличивают поверхность клеток, всасывающих воду, способствуют более быстрому собиранию воды и проведению ее внутрь клеток. Особенно часто развиваются папиллы на клетках у мхов сухих и сильно освещенных мест (тортула - Tortula, барбула - Barbula и др.), но широко представлены и у видов, обитающих в условиях избыточной влажности (у гелодиума шерстистого - Helodium lanatum, аулакомниума болотного - Aulacomnium palustre, у многих лесных мхов из семейства туидиевых -Thuidiaceae, у типичных эпифитов тропических и субтропических лесов из семейства метеориевых - Meteoriaceae).
У ряда мхов проявляется четкое обособление ассимиляционных и водоносных элементов листа. Это характерно для разных систематических групп мхов, далеко отстоящих друг от друга в родственном отношении. Наиболее ярко выраженный диморфизм клеток хорошо известен у сфагновых мхов, а из других листостебельных мхов - только у представителей семейства леукобриевых (Leucobryaceae) и калимперовых (Calymperaceae). Лист состоит из двух типов клеток: из зеленых, содержащих хлоропласты, и из мертвых, бесцветных, лишенных хлоропластов водоносных клеток.
У сфагновых мхов (рис. 44) хлорофиллонооные клетки развиты в большом количестве, окружают водоносные на протяжении всего листа. Водоносные клетки имеют спиральные и кольчатые утолщения и поры - отверстия в наружных оболочках. У леукобриевых в средней части многослойного листа расположено от одного до трех слоев хлорофиллоносных клеток (рис. 45). Почти у всех видов опи со всех сторон окружены водоносными клетками с небольшим количеством пор преимущественно в основании листа.
По экологии и географии две названные группы мхов сильно отличаются. Сфагновые мхи - типичные обитатели болот, особенно широко распространенные в северном полушарии. К леукобриевым в основном относятся эпифиты тропиков и субтропиков, лишь некоторые виды леукобриума (Leucobryum) заходят в северную умеренную зону. Несмотря на это, у тех и у других выработались сходные приспособления в виде водоносных клеток.
Жилка у сфагновых мхов всегда отсутствует. У других верхоплодных мхов большей частью развита простая, т. е. неразветвленная, жилка. У бокоплодных мхов жилка даже в пределах одного семейства может иметь различный характер или отсутствовать. В этом отношении различают наиболее часто встречающуюся простую жилку; двойную, расходящуюся двумя лучами сразу же от основания листа; вильчатую, когда простая, не длинная жилка дает боковые, более короткие ответвления (рис. 43).
В большинстве случаев жилка выступает лишь на нижней, спинной стороне листа (рис. 43, 45), но у некоторых мхов также и на верхней. На спинной стороне жилки часто бывают разнообразные структурные образования (рис. 45). У видов рода фиссиденс (Fissidens) жилка на спинной стороне листа имеет крыловидный вырост. На верхней, брюшной стороне жилки у большинства политриховых находятся продольные, ассимиляционные пластиночки, состоящие из нескольких рядов клеток, богатых хлоропластами (рис. 46). Такие же пластиночки имеются у видов семейства даусониевых (Dawsoniaceae).
В семействе поттиевых у родов алоина (Aloina, рис. 45) и кроссидиум (Crossidium) в верхней части листа на жилке развиваются богатые хлоропластами многоклеточные, иногда разветвленные нити, которые часто подушковидно сплетаются.
Ассимиляционные пластиночки и нити в разных группах выполняют, по-видимому, одни и те же функции: у одних мхов - преимущественно ассимиляционную, у других - обеспечивающую главным образом всасывание воды. При подсыхании у большинства этих мхов края листьев заворачиваются и сверху плотно прикрывают пластиночки и нити. Обламывающиеся нити служат также для вегетативного размножения.
Большинство верхоплодных мхов имеет жилку сложного строения состоящую из указателей, сопроводителей, стереидных пучков и наружных клеток (рис. 43, 45, 46).
Паренхиматические клетки жилки листа с широким просветом, тонкостенные, бедные плазматическим содержанием, расположенные в один, реже в два ряда, называют указателями. Они проводят воду и на продольных стенках нередко снабжены порами. Состояние листа в основном зависит от тургора указателей. В сырую погоду они заполнены водой. Вследствие этого жилка, впитывая влагу, разбухает и вытягивается, а вместе с ней растягивается пластинка листа. В сухую погоду происходит обратное - жилка подсыхает, теряет свое натяжение, сгибается внутрь, а с нею изгибается и пластинка листа.
Сопроводители - тонкостенные, малого диаметра, вытянутые в длину клетки, объединенные в тяж и внешне сходные с клетками простого центрального пучка. Они почти всегда находятся на спинной стороне указателей.
Наружные клетки (обычно с довольно широким просветом) образуют как бы эпидерму жилки.
Стереиды жилки - толстостенные, вытянутые в длину и соединенные в тяжи клетки - сходны с лубяными волокнами сосудистых растений и по своему строению не отличаются от стереидных клеток стебля. Механическая ткань жилки образует один лентовидный стероидный пучок в середине жилки (рис. 45) или у ряда видов - два, спинной и брюшной, разделенные указателями (рис. 43, 45). Только у политриховых и даусониевых жилка имеет большое число близко расположенных стереидных пучков.
Если на все то за 1,5-2,5 ч управлюсь
Время переваривания пищи в желудке, при условии, что она мясная - 4 часа.
Время переваривания пищи в желудке, при условии, что она растительная - 6 часов.
Весь пищеварительный тракт пища проходит в среднем от 12-24 часов.
Ответ:
Симбиониты паразитируют на деревьях, сапротрофы каждый день "разрастаются" все больше, образуют удлинняющийся мицелий, благодаря которому они получают питательные в-ва
От скрещивания петуха с розовидным гребнем с курами, имеющими листовидный гребень, получено 32 цыпленка с листовидным и 27 с роз
ПОМОГИТЕ. Очень срочно нужен не сильно большой реферат-доклад про "ЗАЙЦЕВ"!!Мне надо его сдать 18.01.2014г. Заранее
ОЧЕНЬ нужно, помогите. Спасибо 1.Какая часть органа слуха позвоночных животных развивается только умлекопитающих?1) полость сред
Сформулируйте правила гигиены, которые нужно соблюдать, чтобы не заболеть болезнями, вызываемыми бактериями.
процес сокращения численности осебей, внутресистематических единиц и сужение ареала определенный группы организмов?
Формы приспособления животных и растений в степях, саваннах, пустыне, арктической пустыне и тропического (экваториального) леса.
Листья у мхов сидячие, большей частью поперечно прикрепленные к стеблю, простые, цельные, по краю могут быть зубчатыми и только очень редко глубоко разделенными (рис. 43).
На стебле листья расположены всегда по спирали, образуя двурядное или многорядное листорасположение. Закладываются листья в восходящей последовательности. Рост их начинается с деления двусторонней верхушечной клетки, от которой отделяются сегменты с двух сторон. После прекращения верхушечного роста листья достигают конечного размера посредством вставочного роста, происходящего в их основании.
По расположению на стебле различают листья низовые, срединные (или собственно стеблевые) и покровные.
Покровпые листья окружают гаметангии и обычно отличаются от стеблевых размерами, формой, часто анатомическим строением, а иногда — окраской и консистенцией. Листья, окружающие архегонии, носят название перихециальных, а листья, окружающие антеридии,— перигониальных.
Низовые листья — сильно редуцированные, чешуевидные. Они развиваются в нижней надземной или в подземной части стебля у таких мхов, снабженных ризомом, как климациум (Climacium, табл. 8). Стеблевые листья расположены выше низовых, преимущественно в средней и верхпей частях стебля.

Листовая пластинка чаще однослойная, реже целиком или частично дву-, многослойная. Клетки пластинки обычно богаты хлоропластами и выполняют функцию фотосинтеза. Жилка, если она развита, обычно проходит посередине листа и состоит из толстостенных, вытянутых клеток. Кроме механической роли, она обеспечивает проведение пластических веществ и воды.
По форме листья очень разнообразны (рис. 43). Чрезвычайно различно строение краев листьев. Завернутость и отвернутость краев листа создает микроскопические полости, где и удерживается капиллярная вода. Завернутый край листа, свойственный мхам, обитающим в условиях периодической засухи, выполняет еще и защитную функцию, предохраняя нежные части листа от иссушения.
В листовой пластинке различают клетки двух основных типов: клетки паренхиматические — обычно округлые или угловатые до многоугольных (часто квадратные и шестиугольные), почти равной длины и ширипы, и прозенхиматические — узкие, вытянутые в длину с заостренными, заходящими друг за друга концами.
У мхов вся пластинка листа редко бывает построена из одинаковых клеток. Если верхняя часть листа состоит из паренхиматических мелких клеток, более толстостенных, а иногда мамиллозных или папиллозных, то клетки листового основания почти всегда несколько крупнее и резко отличаются от верхних по форме.
Большой полиморфизм клеток мхов обусловлен не только разнообразием их формы, но и характером их клеточных оболочек. Здесь их многообразие, кажется, не знает пределов.
Кроме разрастания боковых стенок клеток пластинки листа, могут разрастаться и наружные стенки, образуя мамиллозность и папиллозность (рис. 43). Папиллы (утолщения клеточной оболочки) встречаются наиболее часто у бриевых мхов. Они бывают чрезвычайно разнообразной формы, а иногда к тому же в месте их образования клеточная оболочка разрастается и выпячивается. На этом образовании и сидят папиллы. Папиллы сильно увеличивают поверхность клеток, всасывающих воду, способствуют более быстрому собиранию воды и проведению ее внутрь клеток. Особенно часто развиваются папиллы на клетках у мхов сухих и сильно освещенных мест (тортула — Tortula, барбула — Barbula и др.), но широко представлены и у видов, обитающих в условиях избыточной влажности (у гелодиума шерстистого — Helodium lanatum, аулакомниума болотного — Aulacomnium palustre, у многих лесных мхов из семейства туидиевых —Thuidiaceae, у типичных эпифитов тропических и субтропических лесов из семейства метеориевых — Meteoriaceae).
У ряда мхов проявляется четкое обособление ассимиляционных и водоносных элементов листа. Это характерно для разных систематических групп мхов, далеко отстоящих друг от друга в родственном отношении. Наиболее ярко выраженный диморфизм клеток хорошо известен у сфагновых мхов, а из других листостебельных мхов — только у представителей семейства леукобриевых (Leucobryaceae) и калимперовых (Calymperaceae). Лист состоит из двух типов клеток: из зеленых, содержащих хлоропласты, и из мертвых, бесцветных, лишенных хлоропластов водоносных клеток.
У сфагновых мхов (рис. 44) хлорофиллонооные клетки развиты в большом количестве, окружают водоносные на протяжении всего листа. Водоносные клетки имеют спиральные и кольчатые утолщения и поры — отверстия в наружных оболочках. У леукобриевых в средней части многослойного листа расположено от одного до трех слоев хлорофиллоносных клеток (рис. 45). Почти у всех видов опи со всех сторон окружены водоносными клетками с небольшим количеством пор преимущественно в основании листа.
По экологии и географии две названные группы мхов сильно отличаются. Сфагновые мхи — типичные обитатели болот, особенно широко распространенные в северном полушарии. К леукобриевым в основном относятся эпифиты тропиков и субтропиков, лишь некоторые виды леукобриума (Leucobryum) заходят в северную умеренную зону. Несмотря на это, у тех и у других выработались сходные приспособления в виде водоносных клеток.
Жилка у сфагновых мхов всегда отсутствует. У других верхоплодных мхов большей частью развита простая, т. е. неразветвленная, жилка. У бокоплодных мхов жилка даже в пределах одного семейства может иметь различный характер или отсутствовать. В этом отношении различают наиболее часто встречающуюся простую жилку; двойную, расходящуюся двумя лучами сразу же от основания листа; вильчатую, когда простая, не длинная жилка дает боковые, более короткие ответвления (рис. 43).
В большинстве случаев жилка выступает лишь на нижней, спинной стороне листа (рис. 43, 45), но у некоторых мхов также и на верхней. На спинной стороне жилки часто бывают разнообразные структурные образования (рис. 45). У видов рода фиссиденс (Fissidens) жилка на спинной стороне листа имеет крыловидный вырост. На верхней, брюшной стороне жилки у большинства политриховых находятся продольные, ассимиляционные пластиночки, состоящие из нескольких рядов клеток, богатых хлоропластами (рис. 46). Такие же пластиночки имеются у видов семейства даусониевых (Dawsoniaceae).
В семействе поттиевых у родов алоина (Aloina, рис. 45) и кроссидиум (Crossidium) в верхней части листа на жилке развиваются богатые хлоропластами многоклеточные, иногда разветвленные нити, которые часто подушковидно сплетаются.
Ассимиляционные пластиночки и нити в разных группах выполняют, по-видимому, одни и те же функции: у одних мхов — преимущественно ассимиляционную, у других — обеспечивающую главным образом всасывание воды. При подсыхании у большинства этих мхов края листьев заворачиваются и сверху плотно прикрывают пластиночки и нити. Обламывающиеся нити служат также для вегетативного размножения.
Большинство верхоплодных мхов имеет жилку сложного строения состоящую из указателей, сопроводителей, стереидных пучков и наружных клеток (рис. 43, 45, 46).
Паренхиматические клетки жилки листа с широким просветом, тонкостенные, бедные плазматическим содержанием, расположенные в один, реже в два ряда, называют указателями. Они проводят воду и на продольных стенках нередко снабжены порами. Состояние листа в основном зависит от тургора указателей. В сырую погоду они заполнены водой. Вследствие этого жилка, впитывая влагу, разбухает и вытягивается, а вместе с ней растягивается пластинка листа. В сухую погоду происходит обратное — жилка подсыхает, теряет свое натяжение, сгибается внутрь, а с нею изгибается и пластинка листа.
Сопроводители — тонкостенные, малого диаметра, вытянутые в длину клетки, объединенные в тяж и внешне сходные с клетками простого центрального пучка. Они почти всегда находятся на спинной стороне указателей.
Стереиды жилки — толстостенные, вытянутые в длину и соединенные в тяжи клетки — сходны с лубяными волокнами сосудистых растений и по своему строению не отличаются от стереидных клеток стебля. Механическая ткань жилки образует один лентовидный стероидный пучок в середине жилки (рис. 45) или у ряда видов — два, спинной и брюшной, разделенные указателями (рис. 43, 45). Только у политриховых и даусониевых жилка имеет большое число близко расположенных стереидных пучков.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .

Мохообразные растения по количеству видов стоят на первом месте среди нецветковых. Во всем мире их насчитывается 22-27 тыс. видов. На земле мхи встречаются повсеместно. Особенно хорошо они растут в местах, где много влаги. Так, в болотистых местностях они полностью покрывают поверхность земли. Мхи можно увидеть на деревянных строениях, стволах деревьев, поверхности камней и на скалах, и даже в пустынях. Они растут даже на сломанных ветках деревьев, на дне пресных водоемов. В некоторых районах Арктики и Антарктики из высших растений растут только мхи. Раздел ботаники, изучающий моховидные, называется бриологией.
Моховидные — невысокие многолетние, в редких случаях — однолетние травянистые растения. Высота мхов колеблется от 1 мм до нескольких сантиметров, реже — до 60 см (часто погруженные или полупогруженные в воду). У некоторых мхов вегетативные органы не развиты, и тело представлено зеленой пластинкой со спорангием (печёночные мхи) (рис.1, 2-4). Те из мхов, тело которых подразделяется на стебли и листья (филлоиды), называют листостебельные (рис.4, 1). В отличие от водорослей мхи преимущественно сухопутные растения, но предпочитают селиться во влажных местах. Настоящих корней у них нет. Функцию корней выполняют ризоиды (нитевидные выросты). С их помощью мхи поглощают воду и растворенные в ней минеральные вещества, а также прикрепляются к почве.
Рис.1 Многообразие мхов
Стебель у мхов снаружи покрыт оболочкой, состоящей из прозрачных клеток, которые не имеют хлорофилла. Под оболочкой расположена фотосинтезирующая ткань, состоящая из клеток с хлоропластами. У мхов не сформированы сосуды древесины. Поэтому транспортировка воды и минеральных веществ по стеблям осуществляется с участием примитивных, плохо развитых проводящих тканей.
Наиболее широко распространены такие представители моховидных, как зеленый мох (кукушкин лен) и торфяной мох (сфагнум).
Зеленый мох известен также как кукушкин лен. Он относится к группе листостебельных мхов, потому что имеет стебли и листья. Покрывая почву сплошным ковром, кукушкин лен образует травяной пласт, который вытесняет другие зеленые мхи. Он впитывает из атмосферы много влаги, и это вызывает заболачивание почв. Зеленый мох — двудомное растение.
Высота зеленого мха 40 см. Стебель не разветвляется, содержит опорную ткань. Внутри стебля расположены проводящие пучки. К почве прикрепляется с помощью ризоидов.
Питаются зеленые мхи, как и все другие зеленые растения. С помощью ризоидов они поглощают из почвы воду и растворенные в ней минеральные вещества. Листья поглощают углекислый газ воздуха. В результате фотосинтеза образуются органические вещества.
Зеленый мох размножается бесполым и половым путями (рис. 2). Половое и бесполое поколения существуют совместно на одном растении.
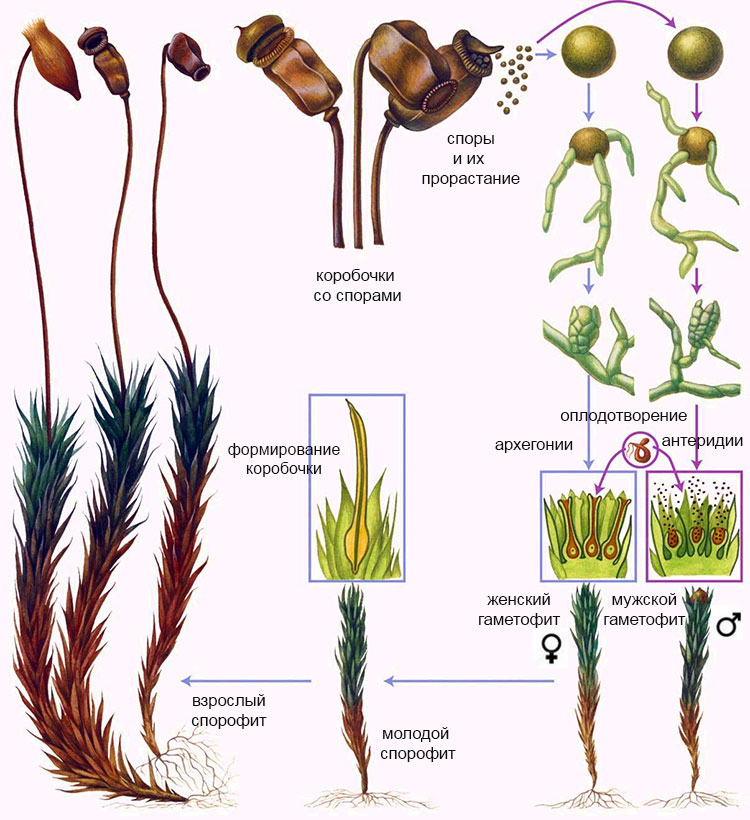
Рис.2 Размножение зеленого мха
При половом размножении образуются мужские и женские гаметофиты. На верхушке мужского растения в антеридиях созревают мужские половые клетки. На верхушке женского растения в архегониях развиваются яйцеклетки. В результате их слияния происходит оплодотворение и образуется зигота. Из зиготы на женском растении (спорофит) развивается коробочка на ножке. В коробочке образуются споры, служащие для бесполого размножения. Созревшие споры высыпаются из коробочки наружу. При благоприятных условиях они прорастают, напоминая сначала разветвленные зеленые водоросли. Из одних проростков развиваются женские растения, из других — мужские.
Торфяной мох, или сфагнум, (рис. 3) — это многолетнее растение с сильно ветвящимся стеблем. В отличие от зеленых и других мхов у торфяных мхов нет не только корней, но и ризоидов. Воду и растворенные в ней минеральные соли поглощают стебли. Светло-зеленые листья сфагнума имеют клетки 2 видов: 1) удлиненные зеленые хлорофиллоносные клетки; 2) водоносные клетки. В зеленых клетках происходит процесс фотосинтеза, и через них от листьев к стеблю проходят органические вещества. Прозрачные мертвые клетки немного крупнее, здесь накапливается запас влаги. У этих водоносных клеток сохранились только оболочки. И когда сфагнум засыхает, т. е. погибают почти все зеленые клетки, листья его кажутся почти белыми. Поскольку сфагнум растет сплошным ковром, он является причиной заболачивания.
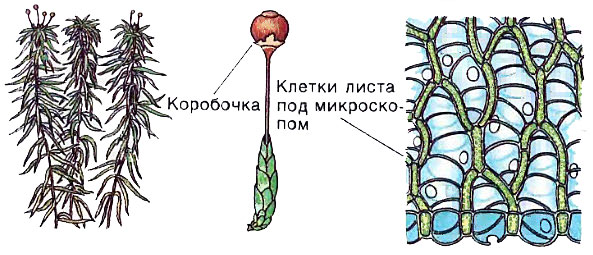
Рис.3 Торфяной мох или сфагнум
Сфагновые мхи — однодомные или двудомные обоеполые растения. Размножаются они половым и бесполым путями. При половом размножении из оплодотворенной яйцеклетки развивается небольшая коробочка, которая содержит множество спор. Бесполое размножение начинается с образования спор. Созревшие споры высыпаются из коробочки наружу. Из них вырастают многоклеточные предростки с многочисленными почками. Именно поэтому мох растет подушками, из которых в будущем развиваются торфяные мхи.
Образование торфа. Торфяные мхи растут в увлажненных местах. Нарастают они верхушками, а снизу отмирают и превращаются в торф. Засохшая часть стеблей из-за нехватки кислорода частично разлагается и образует торфяной слой. Органические кислоты, выделяемые мхом, замедляют его разложение.
Высшие растения — это одно из двух подцарств царства растений. К ним относятся нецветковые и цветковые растения. К высшим нецветковым растениям относятся высшие споровые растения (моховидные, плауновидные, хвощевые и папоротниковидные) и голосеменные.
Моховидные растения имеют стебли и листья. Настоящих корней у них нет. Функцию корней выполняют ризоиды. Широко распространены зеленый мох (кукушкин лен) и торфяной мох (сфагнум).
В природе встречается свыше 300 видов торфяных, или сфагновых, мхов. Торфяные мхи растут на болотах, в тундре, во влажных лесах. У них нет ризоидов. Воду они поглощают с помощью стеблей и листьев. Белыми их называют потому, что большую часть листьев занимает вода, накопленная мертвыми клетками, и цвет листьев становится почти белым. Торфяной мох ежегодно нарастает верхушкой, снизу отмирает и превращается в торф.
Торфяные мхи — основные торфообразователи. Торф — ценное полезное ископаемое. Используется комплексно как топливо, удобрение, теплоизоляционный материал и т. д.
Мхи, как и другие растения, являются необходимым компонентом природы и нуждаются в охране. В Красную книгу Казахстана занесено три вида мхов: сфагнум гладкий, пахифиссинденс крупнолиственный, ортотрихум приглаженный. Одним из мероприятий, направленных на сохранение мхов, является препятствование осушению болот.
Значение мхов. В результате процесса фотосинтеза мхи образуют органические вещества и выделяют в воздух кислород. Растут они на неплодородных почвах и способствуют образованию перегноя (гумуса). Как и другие растения, мхи засыхают и разлагаются при участии микроорганизмов. В результате образуется плодородная торфяная почва, пригодная для произрастания других растений. Мхи являются пищей для многих птиц и используются ими как гнездовой материал.
На торфяных болотах добывают торф, который является горючим полезным ископаемым и широко используется для отопления. Кроме того, он активно используется в качестве строительного материала (для теплоизоляции), удобрения и сырья, а также в химической промышленности. Из торфа получают спирт, карболовую кислоту и т. д.
Мхи — хранилище пресной воды. В заболоченных местностях они принимают участие в сборе воды. Осушение болот зачастую сопровождается исчезновением обитавших на них растений и животных, обмелением некоторых рек, учащением засух.
Мхи — высшие споровые растения (22-27 тыс. видов), живущие на суше во влажных местах. Наука о мхах — бриология. У листостебельных мхов есть стебель, листья, спорангии, вместо корней — ризоиды; у нелистостебельных — только зеленая пластинка 1 со спорангием. У мхов пе развиты сосуды древесины. Мхи фотосинтезируют, выделяют кислород, образуют перегной и торф, заболачивают почву и являются хранилищем 1 пресной воды.
Читайте также: