
Каково значение двойного оплодотворения у цветковых растений
Добавил пользователь Валентин П. Обновлено: 15.09.2024
В процессе эволюции растительного мира у цветковых растений (и только у них) появилось такое явление как двойное оплодотворение, в результате которого образуется семя. У голосеменных растений также образуются семена, но двойного оплодотворения нет. Оплодотворению предшествует опыление, т. е. перенос пыльцы с тычинок одного цветка на пестик чаще всего другого цветка. При двойном оплодотворении в семязачаток проникают два спермия, один из которых сливается с яйцеклеткой, а второй — с крупной центральной клеткой.
Пыльцевые зерна разных цветковых растений имеют различную форму. При этом чаще всего поверхность пыльцевых зерен шероховатая, что позволяет им удерживаться на теле насекомых-опылителей и потом на рыльце пестика. Кроме того, рыльцем выделяется липкая жидкость, удерживающая пыльцу. На рыльце пестика пыльцевое зерно образует пыльцевую трубку, которая растет между клетками рыльца и столбика пестика, после чего врастает в полость завязи пестика.
В полости завязи может находиться один семязачаток, несколько или множество. Их количество зависит от вида растения. Семязачатки по-другому называются семяпочками. Если в завязи несколько семязачатков, то каждый из них опыляется своим пыльцевым зерном (содержащимися в нем спермиями), т. е. в таком случае через пестик будет прорастать несколько пыльцевых трубок.
Семязачатки отрастают от внутренней поверхности стенок завязи в полость завязи. Семязачаток состоит из покрова и ткани центральной части, где образуются восемь гаплоидных клеток (имеющих одинарный набор хромосом). Две из этих клеток сливаются, в результате образуется крупная центральная клетка, у которой восстанавливается двойной набор хромосом.
У семязачатка со стороны, противоположной месту прикрепления к завязи, находится пыльцевход, представляющий собой небольшое отверстие, ведущее к центральной части семязачатка.
В кончике растущей пыльцевой трубки находятся два спермия. У спермиев в отличие от сперматозоидов нет хвостика, и поэтому спермии неподвижны. Когда трубка врастает в семязачаток через пыльцевход, то один спермий сливается с одной из гаплоидных клеток, которая играет роль яйцеклетки. В результате этого оплодотворения образуется зигота с двойным набором хромосом. В последствии из нее развивается зародыш семени.
Второй спермий сливается с центральной клеткой. В результате этого оплодотворения в последствии образуется так называемый эндосперм. Для него характерен тройной набор хромосом, что уникально, так как клетки тела покрытосеменных и многих других организмов имеют двойной набор хромосом.
Эндосперм представляет собой ткань, содержащую запас питательных веществ. Эти вещества зародыш использует в процессе развития семени или при прорастании семени. В первом случае вместо эндосперма в зрелом семени основную массу занимают органы зародыша (чаще всего крупные семядоли), во втором случае — эндосперм остается.
При созревании семени покровы семязачатка превращаются в семенную кожуру.
Двойное оплодотворение у растений имеет большое биологическое значение. Оно было открыто Навашиным в 1898 г. Далее рассмотрим подробнее, как происходит двойное оплодотворение у растений.
Биологическое значение
Процесс двойного оплодотворения способствует активному развитию питательной ткани. В связи с этим семяпочка не запасает вещества впрок. Это, в свою очередь, объясняет ее быстрое развитие.
Схема двойного оплодотворения
Коротко явление можно описать следующим образом. Двойное оплодотворение у покрытосеменных растений состоит в проникновении в завязь двух спермиев. Один сливается с яйцеклеткой. Это способствует началу развития диплоидного зародыша. Второй спермий соединяется с центральной клеткой. В результате формируется триплоидный элемент. Из этой клетки появляется эндосперм. Он является питательным материалом для развивающегося зародыша.
Развитие пыльцевой трубки
Двойное оплодотворение у покрытосеменных начинается после образования гаплоидного сильно редуцированного поколения. Оно представлено гаметофитами. Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.

Формирование клеток: общие сведения
Процесс двойного оплодотворения цветковых растений осуществляется особыми половыми клетками. Их формирование происходит в два этапа. Первая стадия называется спорогенез, вторая – гематогенез. В случае образования мужских клеток эти этапы именуются микроспорогенез и микрогематогенез. При образовании женских половых элементов приставка меняется на "мега" (или "макро"). Спорогенез основывается на мейозе. Это процесс формирования гаплоидных элементов. Мейозу, так же как и у представителей фауны, предшествует размножение клеток посредством митотических делений.
Образование спермиев
Первичное формирование мужских половых элементов осуществляется в особой ткани пыльника. Она называется археспориальной. В ней в результате митозов происходит формирование многочисленных эелементов – материнских клеток пыльцы. Они и вступают затем в мейоз. Вследствие двух мейотических делений образуется 4 гаплоидные микроспоры. Некоторое время они лежат рядом, формируя тетрады. После этого происходит их распад на пыльцевые зерна - отдельные микроспоры. Каждый из образованных элементов начинает покрываться двумя оболочками: внешней (экзина) и внутренней (интина). Затем начинается следующий этап – микрогаметогенез. Он, в свою очередь, состоит из двух митотических последовательных делений. После первого формируется две клетки: генеративная и вегетативная. Впоследствии первая проходит еще одно деление. В результате образуется две мужские клетки – спермии.

Макроспорогенез и мегаспорогенез
В тканях семяпочки начинает обособляться один или несколько археспориальных элементов. Они начинают усиленно расти. Вследствие такой активности они становятся значительно крупнее остальных клеток, окружающих их в семяпочке. Каждый археспориальный элемент один, два или более раз подвергается делению митозом. В некоторых случаях клетка может сразу трансформироваться в материнскую. Внутри нее происходит мейоз. В результате него формируется 4 гаплоидные клетки. Как правило, самая крупная из них начинает развиваться, превращаясь в зародышевый мешок. Три оставшиеся постепенно дегенерируют. На данном этапе макроспорогенез завершается, начинается макрогематогенез. В ходе него происходят митотические деления (у большей части покрытосеменных их три). Цитокинез не сопровождает митозы. В результате трех делений формируется зародышевый мешок с восемью ядрами. Они впоследствии обосабливаются в самостоятельные клетки. Эти элементы распределяются определенным образом по зародышевому мешку. Одна из обособленных клеток, которая, собственно, является яйцеклеткой, совместно с двумя другими – синергидами, занимает место у микропиле, в которое осуществляется проникновение спермиев. В этом процессе синергиды исполняют очень значимую роль. В них содержатся ферменты, которые способствуют растворению оболочек на пыльцевых трубках. В противоположной стороне зародышевого мешка располагаются другие три клетки. Они именуются антиподами. С помощью этих элементов происходит передача из семяпочки питательных веществ в зародышевый мешок. Оставшиеся две клетки располагаются в центральной части. Зачастую они сливаются. В результате их соединения формируется диплоидная центральная клетка. После того как произойдет двойное оплодотворение, и в завязь проникнут спермии, один из них, как выше сказано было, сольется с яйцеклеткой.

Особенности пыльцевой трубки
"Запрет" на самоопыление
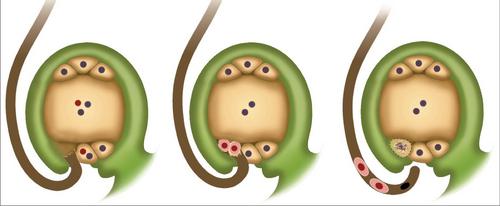
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. "Запрет" на самоопыление проявляется в том, что спорофит "идентифицирует" собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же – короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца – мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.
Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев "самонесовместимость" пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, - s1s2, а вырабатывающего пыльцу - s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.

В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
Развитие управляющих функций мозга ребёнка: полезные советы и упражнения для педагогов
Сертификат и скидка на обучение каждому участнику
ФИО учителя:Жанрыс И.Н
Двойное оплодотворение. Биологическое значение двойного оплодотворения.
Урок ознакомления с новым материалом.
Цели обучения, которые помогает достичь данный урок
описывать значение двойного оплодотворения цветковых растений
Учащиеся будут
понимать и описывать значение процесса двойного оплодотворения цветковых растений.
Критерии оценивания
Учащиеся могут
объяснять механизм и значение двойного оплодотворения
Языковые цели
Учащиеся могут… объяснять механизм двойного оплодотворения цветковых растений
Предметная лексика и терминология :
Генеративные органы растения, цветок, околоцветник, венчик, чашечка, пестик, тычинка, пыльца,раздельнополые цветки, обоеполые цветки, однодомные, двудомные растения, опыление, перекрестное опыление, самоопыление, спорофит, гаметофит, спермий, яйцеклетка, семязачаток, центральная клетка,плод, семя,эндосперм.
Серия полезных фраз для диалога/письма:
Генеративные органы растения –это……
Главными частями цветка являются ……….
Женским органом размножения у цветковых растений является….
Мужским органом размножения у цветковых растений является….
Растения, у которых мужские и женские цветки расположены совместно - ….
Перенос пыльцы с тычинки на рыльце пестика в пределах одного цветка –это…
Продвижение спермиев в семязачаток происходит благодаря росту пыльцевой трубки …
При слиянии спермия с яйцеклеткой образуется зигота, из которой впоследствии развивается зародыш…
Второй спермий сливается с центральной клеткой и образует запас питательных веществ для развития зародыша.
Привитие ценностей
Способствовать формированию навыков сотрудничества, учащихся между собой, уважению друг к другу, природе и экологической грамотности; развивать коммуникативные качества, доверие.
Межпредметные связи
Навыки использования ИКТ
Умение работать с Интернет-ресурсами
Предыдущее обучение
Учащиеся должны знать строение цветка и способы опыления из курса биологии 6 класса
Запланированные этапы урока
Запланированная деятельность на уроке
Приветствие учителя и учащихся друг друга, создание доброжелательной рабочей атмосферы в классе, проверка готовности учащихся к уроку.
Вызов. Актуализация, целеполагание, мотивация.
Проанализируете стихотворение Н. Рыленко
Цветок всю ночь готовит мед,
Пчелу-сластену в гости ждет.
Бери, мол, но как другу, мне окажи услугу.
Пыльцу мучную эту перенеси соседу.
Пчела несет ее, но вот —
Цветок увял и зреет плод.
- Беседа
1. О какой услуге идет речь в стихотворении?
2. Зачем соседу цветка нужна чужая пыльца, ведь у него есть своя?
3. Почему увял цветок?
4. Какой процесс происходит перед тем, как образуется семя и плод?
Формулирование темы и цели урока.
Как происходит оплодотворение у цветковых растений? (при слиянии спермия и яйцеклетки)
Просмотр видео. Рассказ (комментарий) учителя
Двойное оплодотворение — открытый в 1898 г. С. Г. Навашиным половой процесс у цветковых растений, который заключается в том, что один из двух спермиев сливается с яйцеклеткой и образуется зародыш семени, а второй — с центральным ядром зародышевого мешка и в результате образуется эндосперм .
Строение пыльцевого зерна. Прорастание пыльцы. - Объяснение учителя с демонстрацией рисунков, схем, таблиц.
Вопросы учащимся
— Как происходит прорастание пыльцевого зерна?
— Что такое пыльцевая трубка?
Строение семязачатка. Условия двойного оплодотворения у покрытосеменных
- Объяснение учителя с демонстрацией рисунков учебника, схем, таблиц
Сущность двойного оплодотворения у цветковых растений
- Объяснение учителя (демонстрацией рисунков учебника, схем, таблиц)
В результате двойного оплодотворения:
один спермий оплодотворяет яйцеклетку — образуется зигота (2 n );
второй спермий сливается с центральной клеткой — образуется триплоидная клетка (Зn).
В результате образуется:
из завязи — плод;
из семязачатка — семя;
из покрова семязачатка — кожура семени;
из зиготы — зародыш семени (2n);
(ФО) 1. Учащимся необходимо заполнить схему двойного оплодотворения
2. Дополните предложения:
В результате образуется:
из завязи пестика — ________________
из семязачатка — __________________
из покрова семязачатка — ___________
из зиготы — _______________________
Учащийся достиг цели, если:
- правильно заполняет схему оплодотворения:
2 спермий+центральная клетка=эндосперм
- правильно дополняет предложения:
В результате образуется:
из завязи — плод ;
из семязачатка — семя ;
из покрова семязачатка — кожура семени ;
из зиготы — зародыш семени (2n);
1. Что такое оплодотворение? Как происходит двойное оплодотворение у цветковых растений?

Видеоурок способствует формированию представлений о способах опыления цветковых растений, выявляет приспособления к различным способам опыления. Основная цель урока – рассмотреть особенности двойного оплодотворения и его преимущества. В ходе видеоурока подробно рассказывается о процессах микроспорогенеза и макроспорогенеза, их сущности.

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Двойное оплодотворение покрытосеменных растений"
Размножение – это одно из обязательных свойств любого живого организма. Оно заключается в увеличении числа особей.
Различают бесполое и половое размножение растений.
Бесполое размножение подразделяют на спорообразование и вегетативное.
Половое размножение происходит при помощи особых половых клеток –гамет.
При бесполом размножении быстро увеличивается численность вида, все потомки имеют абсолютно такой же генотип, что и родительская особь. А также не происходит увеличения генетического разнообразия, которое может оказаться очень полезным при изменении условий существования вида.
По этой причине большинство живых организмов на Земле размножаются половым путём.
Сущность полового размножения заключается в слиянии генетической информации родителей, благодаря чему генетическое разнообразие в потомстве увеличивается.
У покрытосеменных растений половое размножение связано с цветком.
Важные части цветка – это пестик и тычинка. С их участием происходят сложные процессы полового размножения –опыление и оплодотворение.
Но сперва в будущем цветке начинают образовываться половые клетки.
В жизненном цикле цветковых растений наблюдается смена поколений.
У растений выделяют диплоидное поколение – бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Гаметы образуются в результате митоза, а споры – в результате мейоза. И гаметы, и споры образуются в цветке, поэтому цветок является органом и бесполого, и полового размножения.
Пыльцевое зерно является спорой (микроспорой), а не мужской гаметой, так как в нем самом развиваются мужские гаметы.
У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток.
Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае из 7 клеток (содержит 8 – либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра).
Посмотрим, как образуется мужской гаметофит.
Итак, Микроспорогенез
В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делений становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, поверхность её либо гладкая, либо шероховатая, приспособленная для переноса пыльцы и прилипания её к рыльцу пестика.
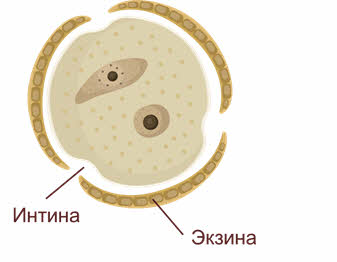
Этим заканчивается микроспорогенез, вслед за образованием одноядерной микроспоры начинается микрогаметогенез.
Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и её ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться ещё в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, называются спермиоклетками, или спермиями.
Таким образом, из одной споры (микроспоры) с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: два из них – спермии и одно – вегетативное. При образовании пыльцевой трубки это вегетативное ядро переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены российским и советским цитологом и эмбриологом растений Сергеем Гавриловичем Навашиным в 1910 г. на лилейных растениях.
После образования гамет пыльник созревает, и пыльца высыпается. Она несёт только генетическую информацию.
Посмотрим, как происходит мегаспорогенез и мегагаметогенез цветковых.
У покрытосеменных растений женский гаметофит – это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез.
В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растёт, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют, судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем её ядро претерпевает ряд делений. При этом сама клетка не делится, а делится только ядро.
У разных систематических групп растений число делений ядра мегаспоры может варьировать от одного до трёх. У большинства растений (70 % видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других – в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
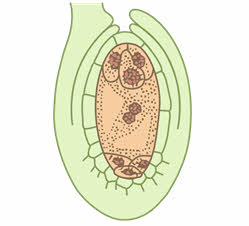
В дальнейшем от каждой из двух полярных четвёрки ядер к центру отходит по одному ядру, которые сливаются, образуя вторичное (центральное) ядро зародышевого мешка. Затем цитоплазма обособляется вокруг ядер гаметофита, который из ядерной стадии развития переходит в клеточную. Три ядра, оставшиеся вблизи халазального полюса, преобразуются в три клетки (антиподы), которые питают гаметофит. Три ядра вблизи микропиле отделяются клеточными перегородками, образуя отдельные клетки: крупную центральную яйцеклетку и две боковые клетки синергиды. Вся цитоплазма, расположенная между антиподами с одной стороны и клетками яйцевого комплекса с другой (яйцеклетка и две синергиды), называется центральной клеткой. В ней находится диплоидное вторичное (центральное) ядро.
На этом этапе женский гаметофит уже полностью сформирован и состоит из шести гаплоидных клеток (одной яйцеклетки, двух синергид и трёх антипод) и одной диплоидной (центральной). Его строение внешне напоминает мешочек, поэтому женский гаметофит покрытосеменных называется зародышевым мешком.
Пылинка попадает на рыльце пестика, и происходит опыление.
Опыление – это перенос пыльцевых зёрен на рыльце пестика, у голосеменных пыльцевые зерна при опылении попадают непосредственно на семязачаток.
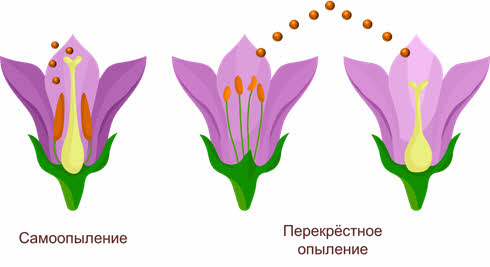
Имеется два основных типа опыления: самоопыление (автогамия) (когда растение опыляется собственной пыльцой) и перекрёстное опыление (аллогамия).
При самоопылении исключён обмен генетической информацией, поскольку пыльцевые зерна попадают на рыльце пестика либо с одной из тычинок этого же цветка, либо с другого цветка, расположенного на том же растении. Это приводит к появлению чистых линий гомозиготных популяций в пределах одного вида, неспособных обмениваться мутировавшими генами, поэтому процессы видообразования в этих популяциях идут самостоятельно.
Перекрёстное опыление – это перенос пыльцы одного растения на рыльце другого. Этот тип опыления встречается более часто, чем самоопыление, между разными особями одного вида происходит обмен аллелями, что приводит к увеличению доли гетерозиготных организмов.
Безусловно, перекрёстное самоопыление имеет большие преимущества по сравнению с самоопылением, поскольку возникшие мутации свободно распространяются в пределах популяции.
Способы опыления у перекрёстноопыляемых цветковых растений весьма разнообразны. Их можно разделить на две группы. Первая: перенос пыльцы осуществляется главным образом насекомыми, а также некоторыми позвоночными (птицами и летучими мышами). Соответственно, различают энтомофилию, орнитофилию и зоофилию. Растения, опыляемые животными, обычно имеют яркоокрашенные крупные цветки. Мелкие цветки, как правило, собраны в соцветия, что зрительно их увеличивает. Для привлечения опылителей служит нектар или большое количество пыльцы, которую опылители охотно поедают.
Вторая группа: пыльца переносится абиотическими факторами – ветром и реже водой, в связи с чем различают анемофилию и гидрофилию.
После попадания пыльцы на рыльце начинается прорастание пыльцевого зерна.
Экзина мужского гаметофита прорывается в области борозды или поры прорастания, и начинает формироваться пыльцевая трубка, которая растёт, проникает в рыльце, через столбик движется по направлению к завязи, где находится семязачаток.
Трубка растёт на кончике. В растущую пыльцевую трубку из пыльцевого зерна перемещается ядро клетки-трубки, а также спермин.
Обычно развитие пыльцевой трубки происходит при уже сформированных семязачатках в завязи. Достигнув завязи, пыльцевая трубка через микропиле проникает внутрь одного из находящихся там семязачатков.
Там пыльцевая трубка направляется к яйцевому аппарату, проникает в одну из синергид и освобождает спермин. Один из спермиев сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами центральной клетки, образуя триплоидное ядро (с тройным набором хромосом). Зигота даёт начало зародышу, а из триплоидной центральной клетки образуется ткань эндосперма.
Таким образом оплодотворение, при котором одна мужская гамета сливается с яйцеклеткой, а вторая ― с вторичным ядром, называется двойным оплодотворением.
Механизм оплодотворения цветковых –двойное оплодотворение, открыто отечественным учёным Сергеем Гавриловичем Навашиным в 1898 году.
Таким образом, оплодотворённый семязачаток развивается в семя, из зиготы возникает зародыш, из триплоидной центральной клетки – эндосперм, а из внешней части семязачатка (интегументов) образуется семенная кожура, из стенок завязи цветка – стенки плода.
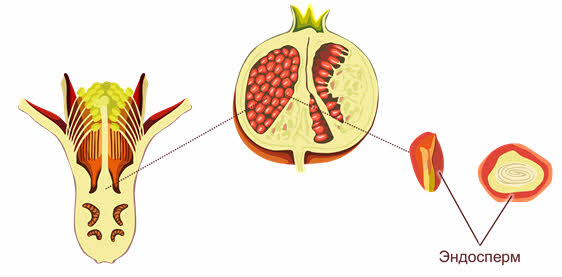
Эндосперм цветковых полностью отличается от первичного эндосперма голосеменных. У них первичный эндосперм представляет собой гаплоидную вегетативную ткань женского гаметофита, где накапливаются питательные вещества семени.
Эндосперм у покрытосеменных происходит из триплоидной центральной клетки, в образовании которой участвуют вторичное ядро женского гаметофита и ядро спермия.
У одних цветковых (например, злаков) эндосперм сильно разрастается и занимает большую часть семени, оттесняя на периферию маленький зародыш.
У других (к пример у бобовых) весь эндосперм поглощается зародышем и используется его семядолями, которые становятся самыми большими структурами зрелого семени.
Читайте также: