
Факторы устойчивости растений к повреждению фитофагами
Обновлено: 08.07.2024
Цель работы – определить взаимодействие систем органов и процессов жизнедеятельности фитофагов с факторами устойчивости растения.
Рекомендации по выполнению
В прил. К показана схема взаимоотношений пшеницы на разных стадиях развития с комплексом фитофагов агроценоза, а также характер повреждений растений. Для выполнения задания нужно изучить прил. К и определить особенности взаимодействия пшеницы с конкретными вредителями. В табл. 4.1 перечислены системы органов, стадии жизнедеятельности, биологические особенности фитофагов, определяющие результаты поиска, заселения растений, откладки яиц, питания личинок и численность потомства вредителей.
Задания выполняются индивидуально, по вариантам. В качестве вариантов приведены виды вредителей пшеницы.
Порядок выполнения задания
1. Покажите в графе 4 взаимосвязь между биологическими особенностями растений, определяющими устойчивость к фитофагам, и биологическими особенностями вредителей (системами органов, жизненными процессами, специализацией). Например, если фитофаг определяет цвет и фазу развития кормового растения с помощью зрения, в графе 4 нужно указать: 1–1, 11–1.
2. В графе 5 укажите, к какой группе факторов устойчивости (антиксенозу, антибиозу или выносливости) принадлежит конкретное биологическое свойство растения. Например, окраска растений относится к группе факторов антиксеноза.
Взаимодействие фитофага с кормовым растением и влияние факторов устойчивости растений на фитофага
| № п/п | Биологические особенности растений | Система органов, стадии жизнедеятельности, специализация фитофага | Взаимодействующие элементы (гр. 2–4) | Фактор устойчивости растений | Влияние фактора устойчивости растения на вредителя |
| 1 | 2 | 3 | 4 | 5 | 6 |
| 1 | Окраска | Зрение | |||
| 2 | Толщина и плотность покровов | Осязание | |||
| 3 | Отложение кремния на поверхности органов | Органы химических чувств | |||
| 4 | Опушенность листьев | Откладка яиц | |||
| 5 | Опушенность колосовых чешуй | Питание взрослых особей | |||
| 6 | Форма колосовых чешуй | Питание личинок | |||
| 7 | Биохимический состав вегетативных органов | Численность потомства | |||
| 8 | Биохимический состав зерна | Физиологические расы | |||
| 9 | Сверхчувствительная реакция клеток | ||||
| 9 | Выполненность соломины | ||||
| 10 | Скорость роста междоузлий | ||||
| 11 | фаза развития растения | ||||
| 12 | Регенерация тканей, органов |
4. Сформулируйте выводы об основных биологических особенностях пшеницы и факторах устойчивости, снижающих вредоносность конкретного фитофага.
Форма предоставления результатов
1. Заполненная табл. 4.1.
2. Выводы об основных факторах, определяющих устойчивость растения к конкретному вредителю.
Варианты
1 – вредная черепашка; 7 – пьявица;
2 – полосатая хлебная блоха; 8 – пшеничный трипс;
3 – шведская муха; 9 – зерновая совка;
4 – гессенская муха; 10 – цветочный пшеничный клещ.
6 – хлебный стеблевой
ТЕМА 5. ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ ИСТОЧНИКОВ
УСТОЙЧИВОСТИ (2 часа)
Цель работы–провести иммунологический анализ устойчивых образцов с помощью тест-культур.
Рекомендации по выполнению
Перед работой изучите раздел 8.4 учебника, посвященный изучению доноров устойчивости для селекции.
Устойчивая реакция сортов дает основания предполагать присутствие у клона гена авирулентности. Устойчивый сорт с геном Pm1 будет защищен от всех клонов возбудителя заболевания, несущих ген А1. Достоверность результатов контролируют, сравнивая реакции набора тест-культур с разными сортами.
Задание выполняется по вариантам, закрепленным за студентом преподавателем. Вариантом служит один из приведенных в табл. 5.2 сортов.
Порядок выполнения задания
1. Определите генотипы всех клонов (тест-культур) мучнистой росы по признаку вирулентности по реакции набора сортов-дифференциаторов. Запишите формулы вирулентности тест-культур по формуле Грина. Например, если клон 2 показывает реакцию R на сорте Аксминстер с геном Pm1, то у клона есть ген авирулентности А1, аналогично определяется присутствие у клона других генов авирулентности.
2. Выделите клоны, поражающие сорт, генотип которого нужно определить. Выпишите их генотипы и выделите общие по отношению к сорту гены вирулентности всех клонов. Эти гены позволят всем клонам преодолевать устойчивость сорта. В генотипе сорта должен содержаться набор генов устойчивости, который может быть преодолен только генами вирулентности, присутствующими у всех клонов, дающих с ним совместимую реакцию.
3. Проверьте полученные результаты, сравнив результаты реакции сорта с предполагаемым генотипом с реакцией клонов, показанной в табл. 5.1.
4. Сделайте вывод о присутствии генов устойчивости у сорта.
© 2014-2022 — Студопедия.Нет — Информационный студенческий ресурс. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав (0.004)
Б.М. Миркин, Л.Г. Наумова
Основы общей экологии
Учебное пособие. М.: Университетская книга, 2005.
– защищаются от фитофагов колючками, образуют розеточные формы с прижатыми к земле листьями, малодоступными для пасущихся животных;
– выделяют запахи, отпугивающие фитофагов.
В то же время эти средства защиты не обеспечивают полную сохранность растений от фитофагов, так как это повлекло бы за собой ряд нежелательных последствий для самих растений:
– несъеденная степная трава превращается в ветошь – войлок, который ухудшает условия жизни растений. Появление обильного войлока ведет к накоплению снега, задержке начала развития растений весной и как итог – к разрушению степной экосистемы. Вместо степных растений (ковылей, типчака) обильно развиваются луговые виды и кустарники. У северной границы степи после этой луговой стадии вообще может восстановиться лес;
– в саванне уменьшение потребления побегов деревьев веткоядными животными (антилопами, жирафами и др.) приводит к тому, что их кроны смыкаются. В итоге учащаются пожары и деревья не успевают восстанавливаться, саванна перерождается в заросли кустарников.
Кроме того, при недостаточном потреблении растений фитофагами не освобождается место для поселения новых поколений растений.
Подобным образом регулируется равновесие между популяциями видов фитопланктона и зоопланктона. Активно поедаемые водоросли быстро размножаются. Некоторые водоросли, напротив, защищаются от выедания специальными выростами на твердых панцирях (как диатомовые) или объединением в большие колонии, которые не могут быть отфильтрованы рачками. Колониальными формами представлено большинство видов цианобактерий. Защите водорослей от выедания помогает их способность образовывать покоящиеся стадии, играющие ту же роль, что и банки семян у растений. Наконец, некоторые водоросли заглатываются планктонными животными‑фитофагами, но не перевариваются и выделяются с экскрементами живыми.
По этой причине невозможно экологическое равновесие, если из экосистемы исчез хищник или в нее внедрился вид‑фитофаг, для которого нет контролирующего его хищника.
При разведении сельскохозяйственных животных плотность популяций фитофагов регулируется человеком, который сам устанавливает норму потребления фитомассы животными. В большинстве случаев при этом норма оказывается завышенной, что ведет к развитию процессов пастбищной дегрессии в экосистемах (см. 12.7).
1. Расскажите об адаптациях, которые позволяют растениям защищаться от фитофагов?


Рис. 1. A — золотарник высочайший (Solidago altissima). B и C — личинка и имаго питающегося на нем золотарникового листоеда (Trirhabda virgata). Изображение из статьи A. Kalske et al., 2019. Insect Herbivory Selects for Volatile-Mediated Plant-Plant Communication
Одна из наиболее явных причин такого переменного во времени и пространстве отбора для популяций растений — сукцессии, то есть закономерные смены сообществ на одной территории (например, смена березняка ельником или зарастание болота). Обычно этот процесс изучается на уровне реакции целого сообщества, но Айно Кальске (Aino Kalske) и Андре Кесслер (André Kessler) из Корнеллского университета (США) решили посмотреть, как популяции одного и того же вида реагируют на изменения пресса со стороны фитофагов на разных стадиях развития травянистого сообщества.
В качестве объекта они выбрали золотарник высочайший (Solidago altissima) (рис. 1, А) — многолетнее сложноцветное, популяции которого живут на заброшенных полях на протяжении десятилетий, переживая изменения в составе окружающей растительности и пресса фитофагов. Кстати, активно разрастающийся аналогичных местообитаниях в России золотарник канадский (Solidago canadensis) иногда рассматривается как синоним этого полиморфного вида.
Согласно гипотезе авторов, слабый пресс со стороны фитофагов, наблюдающийся в этом сообществе на ранних стадиях сукцессии (M. Howard et al., 2018. Eco-evolutionary processes affecting plant–herbivore interactions during early community succession), будет способствовать отбору генотипов с низким уровнем постоянной и индуцированной защиты, затем, на более поздних стадиях, из-за усиления активности фитофагов преимущество перейдет к генотипам с высоким уровнем индуцированной защиты, а если атаки фитофагов не прекратятся и станут более постоянными и предсказуемыми, то отбор будет поддерживать генотипы с высоким уровнем постоянной защиты.
Хотя золотарником питаются более сотни видов членистоногих, наибольший ущерб в месте исследования ему наносят жуки-листоеды Trirhabda virgata (рис. 1, В и С) и Microrhopala vittata. Ранее было показано, что устойчивость к ним наследуется и может эволюционировать в течение одного десятилетия: популяции, не сталкивающиеся с этими фитафагами, теряют устойчивые генотипы (R. F. Bode, A. Kessler, 2012. Herbivore pressure on goldenrod (Solidago altissima L., Asteraceae): its effects on herbivore resistance and vegetative reproduction). Основной способ индуцированной защиты золотарника — выработка в листьях фенольных соединений и дитерпеновых кислот.

Рис. 2. Аэрофотография экспериментального поля, разделенного на участки, находящиеся на разных стадиях девятилетней последовательности: три года засевания кукурузой, и шесть лет зарастания сорной растительностью, в том числе золотарником высочайшим. Два участка (late succession) не засевались 14 и 15 лет. Изображение из дополнительных материалов к обсуждаемой статье в Ecology
В 2017 году исследователи в течение всего сезона развития золотарника каждые три недели учитывали обилие двенадцати видов фитофагов, среди которых были и поедатели листьев, и листовые и стеблевые минеры (прокладывающие внутри тканей растений длинные ходы — мины), и галлобразователи (см. Галл).
Чтобы изучить уровень защиты от фитофагов в золотарниках из разных стадий сукцессии, исследователи собрали от 10 до 15 особей (под особью они понимали совокупность рамет — генетически идентичных стеблей, связанных общим корневищем) на участках, которые претерпели 1, 2, 4, 6 и 14 лет сукцессии после прекращения возделывания кукурузы. Выкопанные золотарники были перенесены в оранжерею, где их корневища дважды дорастили до новых побегов, чтобы избавиться от возможного влияния фитофагов, с которыми растения контактировали в поле.
Анализ сообществ фитофагов показал, что в первые годы сукцессии их состав существенно различается от участка к участку, но к 5–6 году смещается в сторону преобладания наиболее активных потребителей золотарника в регионе, T. virgata и M. vittata (рис. 3). Два участка с 15-летней сукцессией оказались сходны между собой и похожи на часть участков с ранних стадий сукцессии, обилие вышеупомянутых жуков на них также снизилось.
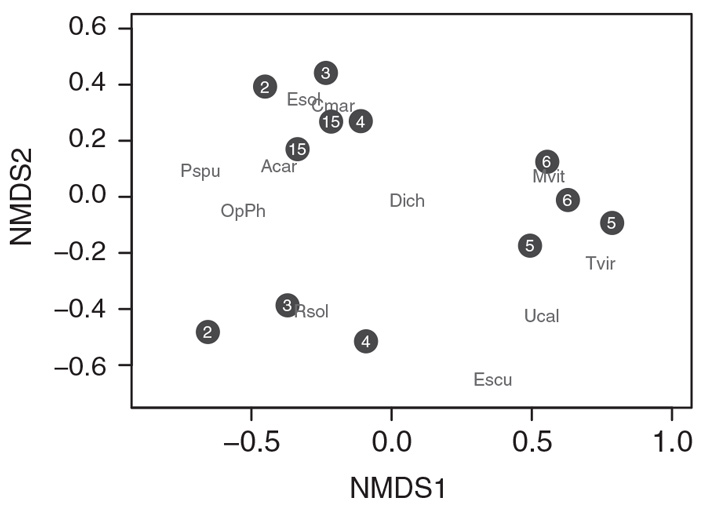
Рис. 3. Сообщества фитофагов на золотарниках, сходство которых изображено с помощью неметрического многомерного шкалирования: чем ближе друг к другу лежат черные кружочки с числами (обозначающими, сколько лет прошло с начала сукцессии), тем более похож состав фитофагов. Буквенные обозначения изображают обилие отдельных видов фитофагов в этом пространстве. Видно, что участки возрастом 5–6 лет похожи друг на друга и в основном населены M. vittata и T. virgata. Обозначения фитофагов: Acar — муха-галлица Asteromyia carbonifera, Cmar — клоп-кружевница Corythucha marmorata, Dich — выемчатокрылые моли Dichomeris spp., Escu — моль-листовертка Epiblema scudderiana, Esol — муха-пестрокрылка Eurosta solidaginis, Mvit — жук-листоед Microrhopala vittata, OpPh — минирующие мухи Ophiomyia sp. и Phytomyza sp., Pspu — пенница Philaenus spumarius, Rsol — муха-галлица Rhopalomyia solidaginis, Tvir — жук-листоед Trirhabda virgata, Ucal — тля Uroleucon caligatum. Рисунок из обсуждаемой статьи в Ecology
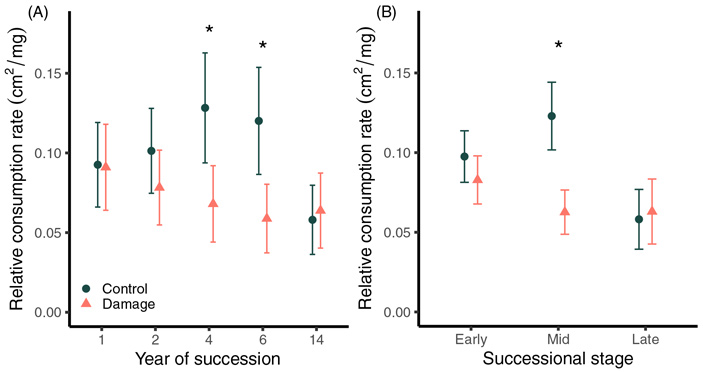
Рис. 4. Относительная скорость потребления золотарниковыми листоедами листвы золотарника высочайшего, взятого с разных стадий сукцессии и подвергшегося (damage) или не подвергшегося (control) предварительно поеданию этим же видом фитофага. Обратите внимание, что скорость потребления обратно зависит от устойчивости золотарника: чем ниже значение по вертикальной оси, тем более устойчив золотарник к поеданию. Звездочками отмечены значимые различия между подвергшимися и не подвергшимися атакам фитофагов золотарниками, свидетельствующие о существовании индуцированной защиты. Возраст участков, с которых были взяты растения, обозначен или как год с начала сукцессии (А), или как стадия сукцессии (В): ранняя, средняя и поздняя. Рисунок из обсуждаемой статьи в Ecology
Также обнаружилось, что скорость роста жуков отрицательно коррелировала с содержанием в листе производных кофейной и кумаровой кислот, но положительно — с содержанием флавоноидов, содержание же производных дитерпеновой кислоты никак не коррелировало с ростом жуков. При этом ни один из исследованных классов веществ не влиял на потребление личинками листьев.
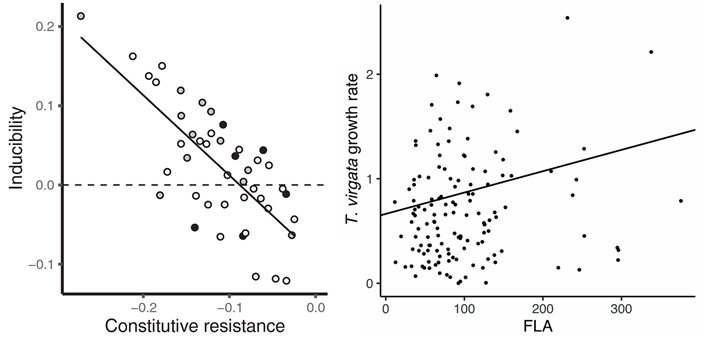
Наконец, в полном согласии с предыдущими исследованиями, постоянная и индуцированная защиты сильно отрицательно коррелировали друг с другом: у золотарников с высоким уровнем одной защиты был низкий уровень другой (рис. 5, слева).
Итак, исследование показало, что наиболее сильный пресс со стороны специализированных фитофагов золотарники испытывают в середине сукцессии, а не в начале и не в конце. Значит ли это, что популяция оказывается наименее устойчивой к поедателям спустя 4–6 лет после возникновения? Скорее всего, ситуация сложнее.
Авторы предполагают следующий сценарий, объясняющий это несоответствие. Где-то после пятого года золотарник перестает возобновляться семенами, так что далее на каждом конкретном участке существуют только определенные клоны. И высокий уровень поедания в средней сукцессии способствовал более активному разрастанию генотипов с высоким уровнем постоянной защиты. Поэтому к поздней сукцессии, когда низкий уровень поедания мог бы снова благоприятствовать индуцированной защите, таких генотипов уже не осталось.
Некоторые из полученных результатов, впрочем, выглядят контринтуитивно. Так, золотарниковый листоед лучше растет на листьях с высоким уровнем флавоноидов, которые вроде как должны защищать от поедания. Авторы предполагают, что дело в том, что T. virgata специализируется на питании растениями этого рода, что сделало его менее чувствительным к защитным веществам своей жертвы (H. V. Cornell, B. A. Hawkins, 2003. Herbivore responses to plant secondary compounds: a test of phytochemical coevolution theory). Впрочем, обнаруженная зависимость не отличается высокой статистической значимостью (р = 0,039), а ее график заставляет предположить, что положительный эффект возникает из-за двух точек в левом верхнем углу (рис. 5, справа).
Обсуждаемая работа не смогла раскрыть всех механизмов, влияющих на изменение проявляемой растениями устойчивости к фитофагам на разных стадиях сукцессии, но смогла показать, что свою роль в этом играет и отбор генотипов с определенной стратегией (в данном случае — с постоянной, а не индуцированной защитой), дающий эффект всего за несколько лет.

Одной из характеристик, выдвигаемых к нынешним сортам агрокультур и технологиям их возделывания, является устойчивость к экологическим воздействиям любой природы возникновения. Чтобы добиться константного результата в динамичных условиях окружающей среды, необходимо не только верно подобрать сорта, а и умело воспользоваться известными техниками по их выращиванию, которые могли бы с наибольшей силой активизировать защитный потенциал растений.
Различные подходы к вопросу защиты растений
Применительно ко многим агрокультурам системная долгосрочная стойкость к стрессовым воздействиям вредителей и экологических факторов все еще представляет собой неразрешенную проблему. И там, где она начинается, появляется вынужденная необходимость применения средств химизации сельского хозяйства, чтобы получить требуемый урожай. Данный вопрос особенно актуален для овощных, фруктовых сортов, широко применимых в пищевой промышленности. А ввиду этого использование пестицидов от фитофагов и возбудителей болезней должно строго лимитироваться.
В то же самое время, ежегодно возрастающая фитосанитарная напряженность во многих странах мира приводит к росту заболеваемости агрокультур, что чревато потерями до 80 % всего урожая. В последние годы на фоне глобальных изменений климата наметилась тенденция к росту убытков растениеводов из-за погодных условий, которые в отдельных регионах планеты приобрели экстремальный характер. Не новость, что климат умеренных широт с умеренно-континентального превратился в резко континентальный, создав новые риски для аграрного производства. Неестественная аридизация (повышение засушливости климата) или, наоборот, гумидизация (увеличение количества осадков и общей влажности) природных условий подбрасывают человечеству все новые и новые задачи, не решить которые означало бы обречь себя на голодное существование.
Все это в комплексе с успехами адаптивно-ландшафтного земледелия, порождает необходимость поисков наиболее экологически оправданных способов вести аграрное хозяйство с наименьшими потерями с одной стороны, и активировать естественную продуктивность агрокультур – с другой.Но решить вышеупомянутые проблемы есть возможность несколькими возможными способами.
Первый из них относится к созданию агросортов, которые были бы устойчивы к воздействию фитопатогенов. В бывших советских республиках – России, Белоруссии, Украине, данная перспектива получила развитие в начале 60-х гг. прошлого века. По сей день этот метод считается одним из наиболее безопасных для естественных экосистем. Однако есть одно весомое но – каждый из конкретных сортов культур способен быть лакомым кусочком для одновременно нескольких десятков вредителей и возбудителей болезней, поэтому придание сортам комплексной стойкости ко всему возможному спектру потенциальных недоброжелателей, к сожалению, практически нереально.
Второй способ связан с широко известными пестицидами. Однако и здесь, кроме рисков для среды обитания и конечного потребителя продукции – человека, имеются множество нюансов. Так, например, культуры, которые дают несколько урожаев на протяжении вегетационного периода, нельзя обрабатывать препаратами, действие которых начинается спустя несколько дней или даже недель. Так способны действовать некоторые фунгициды, проявляя первые признаки своей активности для условий открытых мест высадки лишь спустя 20 суток, а для теплиц – до 7 суток.
Третий путь – это применение биологических средств борьбы. Однако он более применим к защите растений от вредителей, например с использованием энтомофагов или микроорганизмов, вызывающих изменения у конкретных вредоносных насекомых.
Четвертый путь, речь о котором и пойдет дальше в данной статье, связан с повышением иммунного статуса растений через пробуждение естественных защитных факторов, помогающих им справляться с различными воздействиями живой и неживой природы. Данный способ известен уже почти столетие, однако на наших просторах о нем впервые заговорили в начале 70-х годов, начав интенсивные разработки для многих зерновых и овощных культур. Наибольший интерес человечество к данной перспективе наметился лишь в два последних десятилетия, когда кризис в отношениях человека и природы достиг новых масштабов.
Растения и индуцированная устойчивость
Из классической агрономической литературы известно, что приобретенная (индуцированная) устойчивость является ничем иным, как естественной стойкостью растительных организмов, обозначенная их генотипом, которая формируется воздействием массы условий живого и неживого происхождения, отображая степень его приспособительной способности. Она есть не что иное, как непродолжительная устойчивость, обеспечиваемая выраженным действием различных генов, возбуждающих возникновение защитных реакций, почему и представляется как неспецифическая. Активаторами подобной устойчивости может быть широкий спектр веществ абиотического или биотического происхождения.
В отношении заболеваний и их возбудителей индуцированная стойкость растений носит системную направленность. Это проявляется при воздействии вредоносных биологических агентов в течении если не всей, то хотя бы большей части индивидуального развития, что по своим природным характеристикам сильно напоминает природные иммунные ответы у животных.
На сегодняшний день различают два типа индуцированной устойчивости:
системная устойчивость приобретенная (обычно, при перенесении конкретного фитозаболевания) – инфекционный приобретенный иммунитет;
индуцированная системная устойчивость – неинфекционный иммунитет.
Они отличаются между собой по особенностям возникновения. Но связаны необычным на первый взгляд процессом – иммунизацией. В отношении растений это означает процесс, в течение которого происходит активизация естественных механизмов защиты растений. В отличие от приобретенного иммунитета растений, который берет свое начало при успешном преодолении инфекции, что хоть и редко, но все же иногда встречается у растений, данный индуцированный (неинфекционный) иммунитет приобретается в двух случаях:
• при обработке химическими препаратами или же воздействиями физической природы (например, температурными воздействиями, радиоактивным излучением, ультразвуковыми частотами, магнитными полями);
• вакцинационным воздействием, производимым при помощи препаратов биологической природы.
Практический интерес представляют оба способа приобретения индуцированного иммунитета, в то время как практическое значение инфекционного иммунитета не так уж и значимо.
Сегодня индуцированный иммунитет имеет немалое значение в практических агрозащитных мероприятиях. Индуцированный иммунитет, кроме дифференциации по природе индукторов, имеет еще одну классификацию, основанную на характере воздействия. В связи с ней неинфекционный иммунитет бывает локальным или системным. Первое означает, что область его защиты имеет строго ограниченные пределы, например, конкретной вегетативной частью растения. Системный иммунитет не утрачивает своих свойств даже в отдаленных от места поражения растительных компонентах.
Читайте также: