
Наследование окраски плодов тыквы
Добавил пользователь Alex Обновлено: 21.09.2024
Изучая законы Г.Менделя, мы уже знакомились с основными типами взаимодействия аллельных генов. На основе ранее изученного материала заполните таблицу.
Таблица. Типы взаимодействия генов одной аллельной пары
Тип взаимодействия
Характер взаимодействия в
гетерозиготном состоянии
Формулы расщепления по фенотипу в F2 и при анализирующем скрещивании
Примеры
Один аллель (доминантный) полностью подавляет действие другого (рецессивного) аллеля
Высокий рост растения доминирует над карликовостью; вьющиеся волосы – над прямыми; карие глаза – над голубыми и т. п.
Действие каждого аллеля реализуется не полностью, вследствие чего проявляется промежуточный признак
Окраска цветков ночной красавицы; серповидноклеточная анемия; цистонурия и т.п.
Участие обоих аллелей в определении признака
IV группа крови (эритроциты этой группы крови имеют агглютиногены типа А и типа В)
Взаимодействие неаллельных генов
Исходя из законов Г.Менделя, можно прийти к заключению, что существует довольно прочная связь между геном и признаком, что генотип слагается из суммы независимо действующих генов, а фенотип – механическая совокупность отдельных признаков. Однако прямые и однозначные связи гена с признаком скорее исключение, чем правило. Развитие признака организма обычно находится под контролем многих генов, и признак является результатом взаимодействия неаллельных генов.
Простейший случай взаимодействия неаллельных генов – когда признак контролируется двумя парами аллелей. Рассмотрим некоторые примеры такого взаимодействия.
Комплементарное взаимодействие генов
Явление взаимодействия генов, при котором для проявления признака необходимо наличие двух неаллельных генов, называют комплементарностью (от лат. complementum – дополнение), а гены, необходимые для проявления признака, комплементарными, или дополнительными.
Рассмотрим различные варианты комплементарного взаимодействия генов.
• Аллели второго гена проявляются только при наличии доминантного аллеля гена А. Окраска мышей зависит в простейшем случае от двух генов. Мыши с генотипом аа лишены пигмента и имеют белый цвет. При наличии доминантного аллеля А пигмент вырабатывается, и мышь как-то окрашена. Конкретный цвет определяется вторым геном. Его доминантный аллель С обусловливает серый цвет мыши, а рецессивный аллель с – черный цвет. Таким образом, если рассматривать гомозиготные варианты, генотип серых мышей – ААСС, черных – ААсс, белых – ааСС или аасс. При скрещивании серой мыши ААСС с белой аасс в первом поколении все получается по Менделю: все гибриды имеют серый цвет (это гетерозиготы с генотипом АаСс). В F2, как легко проверить, получим 9/16 серых мышей, 3/16 черных и 4/16 белых.
• Для проявления признака в генотипе должны присутствовать доминантные аллели двух разных генов. Примером такого комплементарного взаимодействия генов является наследование формы плода у тыквы. При наличии обоих доминантных аллелей плоды имеют дисковидную форму, при наличии одного (любого!) доминантного аллеля – сферическую, а при отсутствии доминантных аллелей – удлиненную.
• Для проявления признака в генотипе должны присутствовать доминантные аллели двух разных генов, но каждый доминантный аллель в сочетании с рецессивными аллелями другой пары имеет самостоятельное фенотипическое проявление. Например, у кур гороховидная форма гребня определяется одним доминантным геном, розовидная – другим неаллельным ему, но тоже доминантным геном. Когда эти гены окажутся в одном генотипе, развивается ореховидная форма гребня. В случае если организм оказывается гомозиготным по обоим рецессивным генам, развивается простой листовидный гребень. При скрещивании дигибридов (все с ореховидным гребнем) во втором поколении происходит расщепление в отношении 9:3:3:1. Но здесь нельзя найти независимого расщепления каждого аллеля в отношении 3:1, так как во всех случаях совпадения в генотипе обоих доминантных генов их прямого действия не обнаруживается.
Таким образом, о комплементарном взаимодействии неаллельных генов говорят в том случае, когда два неаллельных гена дают новый признак, то есть имеет место новообразование.
Эпистатическое взаимодействие генов
Взаимодействие генов, при котором один ген подавляет действие другого, неаллельного первому, называется эпистазом (от греч. эпи – над и стазис – стоять), а гены, подавляющие действие других генов, называются генами-супрессорами, или генами-ингибиторами (от лат. inhiber – удерживать), или эпистатическими генами. Подавляемый ген называется гипостатическим.
Различают эпистаз доминантный и рецессивный.
• Доминантный эпистаз связан с доминантным геном-ингибитором. Например, ген I обусловливает белый цвет плодов тыквы, при его наличии действие гена В не проявляется. При генотипе iiВВ или iiВb плоды тыквы имеют желтый цвет. Наконец, если оба гена представлены рецессивными аллелями, плоды тыквы имеют зеленый цвет. По этому же типу может наследоваться окраска шерсти собак, овец и других животных.
• Рецессивный эпистаз обнаруживается тогда, когда проявление гена подавляется рецессивными аллелями другого гена. Этот вид эпистаза иллюстрируется наследованием окраски шерсти у домовых мышей. Окраска агути (рыжевато-серая) определяется доминантным геном А, его рецессивный аллель а дает черную окраску. Ген из неаллельной пары В способствует проявлению цветности, а ген b является супрессором и подавляет действие доминантного аллеля А и рецессивного а. Мыши с генотипом Аbb неотличимы по фенотипу от особей с генотипом ааbb – все белые.
Полимерное взаимодействие генов
Полимерия – тип взаимодействия неаллельных генов, при котором несколько пар неаллельных генов действуют на развитие одного признака. Такие гены называются полимерными (от греч. полис – много и мерос – часть). Их обозначают одной буквой, но с разными индексами, которые указывают на число аллельных пар в генотипе, обусловливающих развитие конкретного признака (А1А1А2А2).
Различают два варианта полимерии: с суммирующим действием генов и без усиления генов друг другом.
• Суммирующее действие полимерных генов. В этом случае степень проявления признака зависит от числа доминантных аллелей в генотипе особи. Так, красная окраска зерен пшеницы определяется двумя и более парами генов. Каждый из доминантных генов этих аллелей определяет красную окраску, рецессивные гены определяют белый цвет зерен. Один доминантный аллель дает не очень сильно окрашенные зерна. Если в генотипе присутствуют два доминантных аллеля, интенсивность окраски повышается. Лишь в том случае, когда организм оказывается гомозиготным по всем парам рецессивных генов, зерна не окрашены. Таким образом, при скрещивании дигибридов происходит расщепление в отношении 15 окрашенных к одному белому. Но из 15 окрашенных один будет иметь интенсивный красный цвет, т.к. содержит четыре доминантных аллеля, четыре будут окрашены несколько светлее, т.к. содержат в генотипах три доминантных аллеля и один рецессивный аллель, шесть – еще светлее с двумя доминантными и двумя рецессивными аллелями в генотипах, четыре – еще более светлые, т.к. имеют лишь один доминантный и три рецессивных гена, то есть истинное расщепление будет 1:4:6:4:1.
По данному варианту полимерии наследуются окраска кожи, рост и масса у человека. Подобный же механизм наследования характерен для многих количественных, в том числе и хозяйственно-ценных признаков: содержание сахара в корнеплодах свеклы, содержание витаминов в плодах и овощах, длина колоса злаков, длина початка кукурузы, плодовитость животных, молочность скота, яйценоскость кур и др.
• Полимерные гены не усиливают друг друга. В этом случае расщепление в F2 будет 15:1. Так, плоды пастушьей сумки могут быть треугольными (доминантный признак) и овальными (рецессив). Признак контролируется двумя парами полимерных генов. Если в генотипе растения имеется хотя бы один доминантный аллель из первой или второй пары полимерных генов, то форма плода у него будет треугольной (А1А2; А1а2; а1А2). Овальную форму плодов будут иметь лишь те растения, у которых в генотипе нет ни одного доминантного аллеля – а1а1а2а2.
Таким образом, накопление определенных аллелей в генотипе может привести к изменению выраженности признаков.
Множественное действие генов

Часто ген оказывает действие не на один, а на ряд признаков организма. Явление, при котором один ген может влиять на формирование нескольких признаков организма, называется плейотропией (от греч. плеон – более многочисленный и тропос – поворот).
Такое множественное действие характерно для большинства генов. Однако не следует представлять, что плейотропный ген в равной степени влияет на каждый из признаков. Для абсолютного большинства генов с той или иной степенью плейотропии характерно более сильное влияние на один признак и значительно более слабое – на другой.
Генотип как целостная система
Факт расщепления в потомстве гибридов позволяет утверждать, что генотип слагается из отдельных элементов – генов, которые могут наследоваться независимо (дискретность генотипа). В то же время генотип не может рассматриваться как простая механическая сумма отдельных генов. Генотип – это система взаимодействующих генов. Точнее, взаимодействуют не сами гены (участки ДНК), а образуемые на их основе генные продукты (РНК, а затем – белки). Поэтому в отдельных случаях действие разных генов относительно независимо, но, как правило, проявление признака есть результат взаимодействия продуктов разных генов.
Генотип любого организма представляет собой сложную целостную систему взаимодействующих генов. Эта целостность генотипа возникла исторически в процессе эволюции вида. В результате мутаций постоянно появляются новые гены, формируются новые хромосомы и даже новые геномы. Вновь возникшие гены могут сразу же взаимодействовать с уже имеющимися генами или модифицировать характер работы последних, даже будучи рецессивными, то есть, не проявляясь сами по себе.
Следовательно, у каждого вида растений и животных генотип проявляет себя как исторически сложившаяся к данному моменту целостная система.
II. Закрепление знаний
Обобщающая беседа по ходу изучения нового материала.
III. Домашнее задание
Изучить параграф учебника (связь между генами и признаками, типы взаимодействия аллельных и неаллельных генов, плейотропия, генотип как целостная система).
Свидетельство и скидка на обучение каждому участнику
Выберите документ из архива для просмотра:
Выбранный для просмотра документ Презентация 10 класс неаллельные гены.pptx
Описание презентации по отдельным слайдам:

Комплементарность – это тип взаимодействия генов, который возникает в случае, когда за развитие одного признака отвечают два гена, и действие одного гена всегда дополняется действием другого гена





Эпистаз – это тип взаимодействия генов, который возникает в случае, когда аллели одного гена подавляют действие аллелей другого гена; это могут быть доминантные, рецессивные, либо и доминантные и рецессивные аллели


Пример № 2. Наследование окраски плодов у тыкв: В – желтая окраска; b – зеленая окраска; А – подавляет проявление окраски; а – не препятствует проявлению окраски; В-А- – белая окраска; bbA- – белая окраска; В-аа – желтая окраска; bbaa – зеленая окраска плодов.

Пример № 3: Наследование окраски оперения у кур: А – черная окраска; А – белая окраска; I – ген, подавляющий проявление окраски; i – ген, не препятствующий проявлению окраски; А-I- – белая окраска; ааI- – белая окраска; А-ii – черная окраска

Полимерия – это тип взаимодействия генов, когда за развитие одного признака отвечают два или несколько генов однозначного действия


Р ♀ A1A1А2А2 ♂ a1a1а2а2 треугольная округлая Г A1А2 a1а2 F1 A1a1А2а2 треугольные F2 A1А1А2А2, А1 А1 a2a2, a1a1А2А2; а1а1а2а2 треугольные округлые 15/16 1/16 Некумулятивная полимерия на примере плодов пастушьей сумки

Пастушья сумка обыкновенная Capsella bursa-pastoris (L.)

Р А1А1А2А2 а1а1а2а2 негр белый F1 16 А1а1А2а2 Мулаты F2 А1А1А2А2 4А1А1А2а2, а1а1а2а2 негр 6А1а1А2а2 БЕЛЫЙ 4А1а1а2а2 мулаты 1/16 14/16 1/16 негр мулаты белые

Плейотропия возникает в том случае, если один ген определяет развитие нескольких признаков и свойств организмов

Синдром Марфана – пример проявления плейотропии

Никколо Паганини (27.10.1782-27.05.1840) итальянский скрипач-виртуоз, гитарист, композитор

ВЫВОДЫ: 1. Изучение неаллельных типов взаимодействия генов имеет важное значение в понимании закономерностей наследования признаков. 2. Формирование признаков в ходе индивидуального развития организма обусловливается его генотипом и влиянием внешней среды. 3. Взаимодействие неаллельных генов приводит к появлению необычных соотношений фенотипов потомков во втором поколении при дигибридном скрещивании.
Выбранный для просмотра документ картинки.docx






Паучьи пальцы у человека

Выбранный для просмотра документ конспект урока.docx
Тема: ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Учебник: Биология. Общая биология 10–11 классы: учебник для общеобразовательных учреждений / А. А. Каменский, Е. А. Криксунов, В. В. Пасечник.- М.: Дрофа, 2010.
Тип урока : изучение нового материала (объяснение темы урока идет с использованием презентации).
Цели урока :
Образовательные: сформировать знания о взаимодействии неаллельных генов (комплементарность, эпистаз, полимерия);
Развивающие : развивать представления о закономерностях наследования признаков; продолжать развивать умения пользоваться терминами, символами генетики; продолжить формирование навыков решения генетических задач;
Воспитательные : формирование бережного отношения к своему организму как носителю уникального генотипа; развивать представления о влиянии окружающей среды на формирование фенотипа.
Оборудование и материалы:
1) проектор, переносной экран, компьютер;
3) дополнительная информация о синдроме Марфана, задачи
Использованные источники:
Сборник задач по молекулярной биологии и генетике: учебное пособие / В. В. Дубейковская. – Тирасполь: 2000.
1.Организационный момент.
“ Разуму свойственно размышлять, то есть связывать причины и следствия, давать ответ на вопрос “почему” и находить в цепи происходящего начало и конец”. Ж. Фабр.
“ Пока живёшь, обучайся.
Не жди, когда старость принесет тебе мудрость”
Учитель: Наш урок пройдет под девизом: “Думать коллективно, работать оперативно, спорить доказательно – для всех обязательно”.
2. Разминка. Работа с индивидуальными карточками по основным терминам темы (на 3 варианта).
Генотип, фенотип, ген, 3 закона Г.Менделя
3. Изучение новой темы.
Проблема урока
Почему у потомства иногда появляются признаки не свойственные их родителям?
Данный урок даёт нам возможность “приоткрыть дверь” научного познания процессов, понять взаимосвязь процессов в организме через различные виды взаимодействия генов в генотипе.
Учитель дифференцирует на группы обучающихся по форме учебных действий. Работа 2 и 3 группы направлена на самостоятельную работу по углубленному изучению материала, решению задач повышенной сложности. Дети 1-й группы, т.е. школьники с низким уровнем обучаемости, выполняют задания под руководством учителя.
Учитель: развитие каждого признака обеспечивается всем генотипом, и все гены в генотипе влияют на развитие каждого признака. Также бывают случаи, когда один ген влияет на формирование нескольких признаков. Все эти механизмы наследования объясняются закономерностями, установленными Г. Менделем. Давайте вспомним, какая общая формула расщепления по фенотипу используется при взаимодействии двух генов для F 2 ?
Учащиеся : 9 / 16 А-В- : 3 / 16 А-вв : 3 / 16 ааВ- : 1 / 16 аавв.
Учитель: для анализирующего скрещивания расщепление в F 1 ?
Учащиеся : ¼ А-В- : ¼ А-вв : ¼ ааВ- : 1 / 4 аавв.
Учитель: характерной особенностью при взаимодействии неаллельных генов является развитие в потомстве новых вариаций признаков. Менделевские расщепления модифицируются, т. к. некоторые классы по фенотипу не отличаются друг от друга, их число суммируется. Так, при взаимодействии двух генов вместо расщепления 9:3:3:1 может быть 12:3:1, 9:7, 9:6:1, 9:3:4 и т. д. В классической генетике наиболее изученными являются следующие взаимодействия неаллельных генов: комплементарность, эпистаз, полимерия. Они определяют признаки: цвет кожи, волос, радужной оболочки глаз, глухота, рост у человека, окраску животных, зерен пшеницы, морфологические признаки животных и т. д.
Все это поможет нам решать задачи на взаимодействие генов и использовать те знания, которые мы получили с вами до этого.
Первый тип взаимодействия неаллельных генов носит название комплементарность. Это тип взаимоотношения доминантных неаллельных генов, которые при совместном нахождении в генотипе обусловливают проявление нового признака. Наблюдаются различные классы фенотипических расщеплений.
Например, наследование форм гребня у кур, при котором наблюдается фенотипическое расщепление 9:3:3:1 .
А – розовидный гребень А- bb – розовидный гребень
а – нормальный гребень аа B - - гороховдный гребень
В – гороховидный гребень А-В- - ореховидный гребень
b - нормальный гребень aabb – листовидный (нормальный) гребень
Расщепление в соотношении 9:6:1 наблюдается у тыквы, формы плода которой могут быть сферическими, дисковидными и удлиненными
А или В – сферическая форма (9)
А-В- дисковидная форма (6)
аа bb – удлиненная форма (1)
Также примерами комплементарного взаимодействия может быть: окраска меха у норки, мышей, окраска глаз у дрозофилы.
Эпистаз – тип взаимодействия неаллельных генов, при котором аллели одного гена подавляют действие аллелей другого (ему не аллельного).
Гены-подавители называются ингибиторами или супрессорами. Если ингибитор доминантен, то и эпистаз называется доминантным. При рецессивном ингибиторе эпистаз - рецессивный. В случае доминантного эпистаза могут быть два типа фенотипического расщепления:
а) расщепление 12:3:1 .
Например: у тыквы окраска плода может быть желтой ( А ) и зеленой ( а ). Эта окраска может подавляться доминантным ингибитором ( I ), в результате чего появляются белые плоды:
9 I- А- : 3 I- аа : 3 ii А- : 1 iiаа А – желтые плоды а – зеленые плоды тыквы
белые желтые зеленые I – подавляет окрашивание i – не подавляет окрашивание
б) расщепление 13:3
Например: у лука окраска кожуры может быть ( А ) и нет окрашивания ( а ), причем пигмент может подавляться доминантным ингибитором ( I) или не подавляться его рецессивным аллелем ( i ):
9 I- А- : 3 I- аа : 1 iiаа : 3 ii А-
Еще одним примером взаимодействия неаллельных генов может быть полимерное действие генов . Полимерия – когда за 1 признак отвечает несколько неаллельных генов. Чтобы подчеркнуть их однозначность, одинаковость действия генов, их обозначают одними и теми же символами, добавляя лишь разные цифры: А 1 , А 2 и т. п. Существует два вида полимерии:
а) кумулятивная - степень выражения признака зависит от числа доминантных генов. По этому типу наследуются все количественные признаки, которые можно измерить и подсчитать.
Например: окраска зерен пшеницы или интенсивность окраски кожи человека за счет пигмента меланина (приложение 3). В таких случаях наблюдается расщепление по фенотипу в соотношении
В этом случае характер проявления признака не меняется в зависимости от числа доминантных генов в генотипе, для его проявления достаточно 1 доминантного аллеля любого гена.
Например: у растения пастушья сумка треугольная форма плода определяется доминантными аллелями разных генов – А 1 и А 2 , а рецессивные аллели а 1 и а 2 определяют овальную форму плода :
Плейотропность - это такое взаимодействие, при котором один ген контролирует развитие нескольких признаков. Это может быть связано с тем, что один ген отвечает за формирование фермента, который влияет не только на свою реакцию, но также оказывает влияние на вторичные реакции биосинтеза. Пример – синдром Марфана.
Мутантный ген определяет нарушение развития соединительной ткани (вывих хрусталика глаза, пороки клапана сердца, пороки развития сосудов, частые вывихи суставов, длинные и тонкие пальцы, т.е. приводит к развитию Синдрома Марфана.
4. Закрепление новых знаний.
Чтобы закрепить данный материал, попробуйте решить задачи , приложение 3 ).
5. Подведение итогов.
Учитель: мы узнали, что генотип – это не простая совокупность генов, а система сложного взаимодействия между ними. Какие же выводы из сегодняшнего урока можно сделать?
Учащиеся делают выводы с помощью учителя и проверяют правильность своих умозаключений:
1. Изучение неаллельных типов взаимодействия генов имеет важное значение в понимании закономерностей наследования признаков.
2. Формирование признаков в ходе индивидуального развития организма обусловливается его генотипом и влиянием внешней среды.
3. Взаимодействие неаллельных генов приводит к появлению необычных соотношений фенотипов потомков во втором поколении при дигибридном скрещивании.
6. Рефлексия.
Что нового, интересного вы узнали на сегодняшнем уроке?
6. Заключительный этап урока:
а) домашнее задание: § 43, решить задачу;
У мухи дрозофилы наряду с дикой красной окраской глаз (А-В-) встречается коричневая (А-вв), ярко-красная (ааВ-) и белая (аавв). Определить расщепление по фенотипу при скрещивании двух гетерозиготных особей по анализируемым генам.
б) выставление оценок и анализ активности учащихся на уроке.
Приложение 1.
Единственная компенсация, которую люди с синдромом Марфана получают от судьбы за свой порок, - повышенное содержание адреналина в крови. Как известно, этот гормон вырабатывается надпочечниками и выбрасывается в кровяное русло в момент опасности. В результате многие параметры человеческого организма (сердцебиение, давление крови) приводятся, так сказать, в боевую готовность. Таким образом, люди с синдромом Марфана всю жизнь находятся в возбужденном состоянии: адреналин постоянно подстегивает нервную систему и делает их невероятными трудоголиками.
Исторические примеры: известность и синдром Марфана
Нервное напряжение, в котором, по-видимому, постоянно находился этот талантливый человек, порождало у него множество страхов. Он боялся заболеть холерой, пострадать от пожара, попасть в аварию, потерять важные документы, принять не ту дозу лекарства. История знает случай, когда длинные, тонкие пальцы человека с синдромом Марфана вместе с впечатляющей работоспособностью помогли их обладателю сделать
фантастическую карьеру. Речь идет о знаменитом скрипаче Никколо Паганини. Гете и Бальзак так описывают его внешность в своих воспоминаниях: мертвенно-бледное, как будто вылепленное из воска лицо, глубоко запавшие глаза, худоба, угловатые движения и, самое главное, тонкие сверхгибкие пальцы, какой-то невероятной длины, как будто вдвое длиннее, чем у обычных людей. Эта чисто морфологическая особенность позволяла ему творить со скрипкой настоящие чудеса. В толпе, слушавшей импровизации Паганини на римских улицах, одни говорили, что он в сговоре с дьяволом, другие - что его искусство является музыкой небес, в которой звучат ангельские голоса. Он играл так, что слушателям казалось, будто где-то спрятана вторая скрипка, играющая одновременно с первой. Многие вплоть до XX в. верили слухам, что в молодости Никколо прибег к помощи хирурга, который сделал ему операцию, чтобы повысить гибкость рук. Теперь-то мы знаем, что, скорее всего, своими данными он был обязан редкому генетическому отклонению.
Из наших современников синдромом Марфана, возможно, страдал биолог Г. В. Никольский. Ко времени окончания Московского университета он имел уже пять печатных трудов. За 30 последующих лет работы число его печатных публикаций превысило 300, причем среди них было около десяти книг. Такой потрясающей работоспособностью может похвастаться далеко не каждый даже очень способный ученый! Можно ли после этого утверждать, что любые обусловленные генами нарушения в развитии являются, безусловно, вредными?
Приложение 2.
1. Бежевая норка скрещена с серой. В F 1 все норки коричневые. В F 2 получилось 14 серых, 46 коричневых, 16 белых, 5 кремовых норок. Как наследуется признак?
2. Цвет кожи человека определяется взаимодействием нескольких пар генов по типу полимерии, т.е. цвет кожи тем темнее, чем больше доминантных генов в генотипе. Возможные генотипы и фенотипы цвета кожи:
черная кожа – A 1 A 1 A 2 A 2
темная – A 1 A 1 A 2 a 2
смуглая ( мулат ) – A 1 a 1 A 2 a 2
светлая – A 1 a 1 a 2 a 2
белая – a 1 a 1 a 2 a 2
Если два мулата ( A 1 a 1 A 2 a 2 ) имеют детей, то можно ли ожидать среди них детей с черной, смуглой и белой кожей? Какую часть составят дети каждого типа?
Например, у кур ореховидная форма гребешка вызвана взаимодействием доминантных аллелей комплементарных генов А и В (А_В_). Если же сочетаются один ген в доминантном состоянии, а другого в рецессивном, то наблюдается появление или розовидного гребешка (А_bb), или гороховидного (ааВ_). Те куры, у которых образовался генотип aabb, имеют гребешок листовидный. В пропорциональном выражении это выглядит так: ореховидная форма — 9/16, розовидная и гороховидная по 3/16, и минимум выпадает на листовидную —


1/16. Расщепление по фенотипу 9:3:3:1.

Добавим, что при комплементарности расщепление по фенотипу может быть не только такое, как в примерах, приведенных выше (у кур 9:3:3:1, у баклажанов 9:7), но и другое — 9:6:1 или 9:3:4.
Эпистатичные гены
Эпистаз — такое взаимодействие неаллельных генов, при котором активность одного гена — эпистатичного, подавляет активность другого — гипостатичного.
Эпистатичные гены также называют ингибиторами или супрессорами, то есть подавляющими задерживающими — их буквенное обозначение I (i).
Эпистаз связан с синтезом регуляторных белков на основе регуляторного гена. Есть два случая.
1. Регуляторный ген активен, синтезирует белок-репрессор, препятствующий транскрипции основного структурного гена, определяющего признак.
2. Регуляторный ген не способен синтезировать белок-активатор, который будет регулировать активность структурного гена. По сути, регуляторный ген нарушен.
3. В обоих случая не синтезируется белок, отвечающий за признак.
Если эпистатичный ген доминирует, эпистаз тоже доминантный. Расщепление по фенотипу в этом случае может быть 12:3:1, 13:3 или 7:6:3. Пример доминантного эпистаза — передача потомству белой окраски плодов у тыквы. Допустим, ген W задает появление белых тыквин, а рецессивный ген w подавляет. Ген Y ведает желтой окраской, ген y зеленой. Если скрестить белые тыквы с зелеными, весь урожай будет белым, так как ген W подавляет Y и y. В результате белых плодов — 12/16, желтых — 3/16, зеленых — 1/16. Расщепление по фенотипу здесь 12:3:1.
Когда эпистатичный ген рецессивный — эпистаз тоже рецессивный и идет расщепление по фенотипу 9:3:4, 9:7 или 13:3. Примером здесь может служить наследование мышами белой шерстки. В представленной схеме ген А ответственен за появление серой шерсти, ген а — черной, I — не влияет на проявление признака, i — ген эпистатичный, который подавляет гены.
Когда эпистатичный ген рецессивный — эпистаз тоже рецессивный и идет расщепление по фенотипу 9:3:4, 9:7 или 13:3. Примером здесь может служить наследование мышами белой шерстки. В представленной схеме ген А ответственен за появление серой шерсти, ген а — черной, I — не влияет на проявление признака, i — ген эпистатичный, который подавляет гены.


А и а и отвечает за белую окраску (мыши-альбиносы). Пропорция такова: серая окраска — 9/16, черная — 3/16, белая — 4/16. Расщепление по фенотипу 9:3:4.
1. При кумулятивном действии генов сила проявления признака зависит от суммы их действий — чем выше количество доминантных аллелей, тем ярче выражен признак. Кумулятивная полимерия наблюдается при наследовании, например, цвета кожи человеком. Количество в коже меланина определяют три неаллельных гена: А1, А2, А3.
2. При некумулятивном действии не важно количество доминантных аллелей — даже при наличии одного из доминантных аллелей признак проявится. Такая полимерия наблюдается, например, у растения пастушья сумка в наследовании формы плодов: треугольные плоды — 15/16, овальные — 1/16.
Плейотропией называют зависимость проявления нескольких фенотипических признаков от действия одного гена. Здесь властвует биохимия — белок (фермент), контролируемый определенным геном, задает развитие некоего признака и одновременно воздействует на вторичные реакции биосинтеза нескольких прочих признаков.
Еще Грегором Менделем было замечено, что растения гороха с темно-красными цветками дополнительно всегда имели выраженные розовые пятна в пазухах листьев и к тому же серую семенную кожуру. А у дрозофил, например, белоглазие связано с низкой плодовитостью и короткой продолжительностью жизни. Выходит, один наследственный фактор (ген) влиял на все эти признаки. Механизмы плейотропии изучали Томас Морган, Сергей Четвериков, Николай Вавилов. Плейотропия распространена повсеместно, вероятно, плейотропный эффект присущ большинству генов. Она бывает первичная и вторичная.
При первичной плейотропии ген проявляет множественное действие одновременно. Пример такой работы гена — синдром Марфана, при котором наблюдается очень высокий рост, тонкие и чересчур длинные конечности и пальцы, искривление грудины, плоскостопие, аневризма аорты, смещение хрусталика и другие патологии. Причина синдрома Марфана в доминантной мутации гена, который одновременно отвечает за рост человека, длину пальцев, местоположение и форму хрусталика и пр.
Признаки, наследование которых мы рассматривали ранее, контролируются генами одной аллельной пары. Однако многие признаки живых организмов определяются взаимодействием двух или более пар генов, т. е. неаллельными генами. Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия.
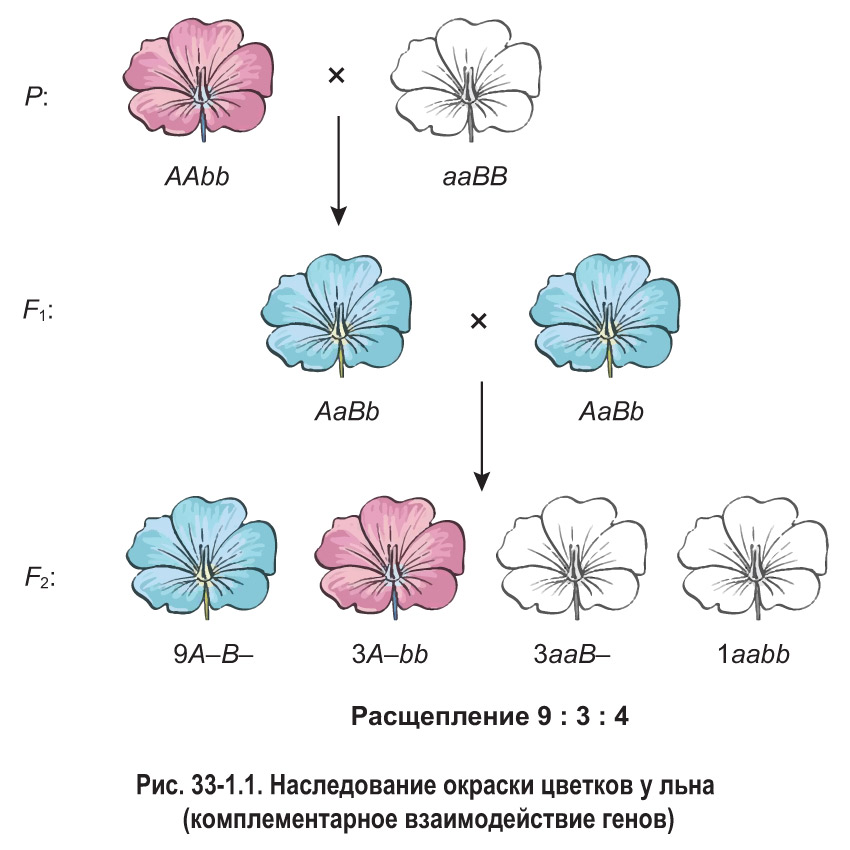
Комплементарность выражается в том, что доминантные гены разных аллелей совместно обусловливают появление нового признака, не определявшегося ни одним из этих генов по отдельности. Например, у льна доминантный ген одной аллели (А) определяет розовую окраску цветков, а доминантный ген другой аллели (В) — белую. Если в генотипе растения присутствуют оба доминантных аллеля, его цветки имеют голубую окраску. Таким образом, неаллельные гены А и В действуют комплементарно (взаимно дополняя друг друга), что приводит к формированию нового признака. Суть такого взаимодействия можно отразить в виде схемы.

Как видно из схемы, синтез голубого пигмента из неокрашенного предшественника (пропигмента) осуществляется в два этапа, каждый из которых катализируется особым ферментом. Присутствие этих ферментов в клетках лепестков льна определяется наличием в генотипе соответствующих генов — А и В. Так, ген А определяет наличие фермента, который превращает пропигмент в промежуточный продукт — пигмент розового цвета. Ген В обусловливает присутствие второго фермента, синтезирующего голубой пигмент из розового. Поэтому у растений, имеющих оба доминантных аллеля, в лепестках образуется конечный продукт — пигмент голубого цвета. Если отсутствует ген В (а значит, и соответствующий фермент), в клетках протекает лишь первая стадия — образование розового пигмента. В случае отсутствия гена А (или обоих доминантных аллелей) не осуществляется даже первый этап синтеза пигмента, и лепестки остаются белыми.
Рассмотрим скрещивание дигомозиготных растений льна, имеющих розовые (AAbb) и белые (aaBB) цветки (рис. 33-1.1). В первом поколении наблюдается единообразие гибридов. Все они имеют голубые цветки, что обусловлено наличием в генотипе как гена А, так и гена В. Во втором гибридном поколении происходит расщепление в соотношении 9 голубых : 3 розовых : 4 белых.
Комплементарное взаимодействие генов лежит в основе проявления ряда признаков у различных организмов. Так, по типу комплементарности взаимодействуют гены, контролирующие окраску коконов у тутового шелкопряда и цветков у душистого горошка, форму гребня у кур, цвет оперения у попугаев и т. д. В зависимости от особенностей фенотипического проявления генов в потомстве от скрещивания дигетерозигот могут наблюдаться различные варианты расщепления — 9 : 7, 9 : 3 : 4, 9 : 6 : 1, 9 : 3 : 3 : 1.
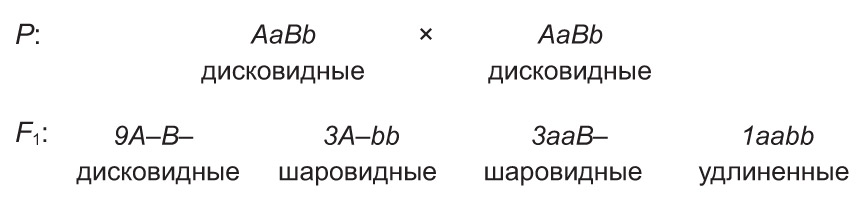
Например, у тыквы доминантные аллели А и В совместно приводят к появлению плодов дисковидной формы, а по отдельности — шаровидной. В то же время рецессивные аллели этих генов обусловливают удлиненную форму плодов. При скрещивании дигетерозиготных растений, имеющих дисковидные плоды, в потомстве происходит расщепление 9 : 6 : 1.
Сходным образом наследуется цвет оперения волнистых попугаев. Доминантный ген одной аллели (А) обусловливает у них голубую окраску перьев, а доминантный ген другой аллели (В) — желтую. При наличии в генотипе обоих доминантных аллелей (А и В) проявляется новый признак — зеленое оперение. Вместе с тем рецессивные аллели этих генов (a и b) определяют белую окраску перьев. Поэтому скрещивание зеленых дигетерозиготных особей приводит к расщеплению по цвету оперения среди потомков в соотношении 9 : 3 : 3 : 1.
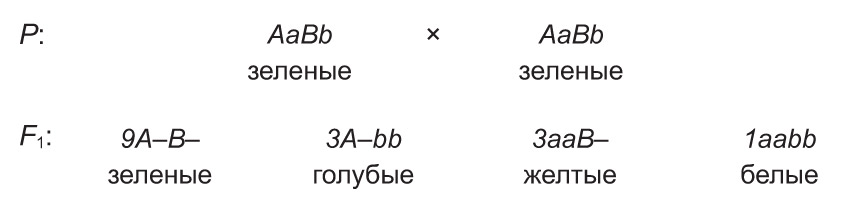
Эпистаз — тип взаимодействия, при котором ген одной аллели подавляет проявление генов другой аллельной пары. Такие гены, блокирующие фенотипическое проявление других неаллельных генов, называются эпистатическими, генами-ингибиторами или супрессорами. Подавляемые гены, в свою очередь, получили название гипостатических. В зависимости от того, доминантным или рецессивным является ген-ингибитор, различают доминантный и рецессивный эпистаз.
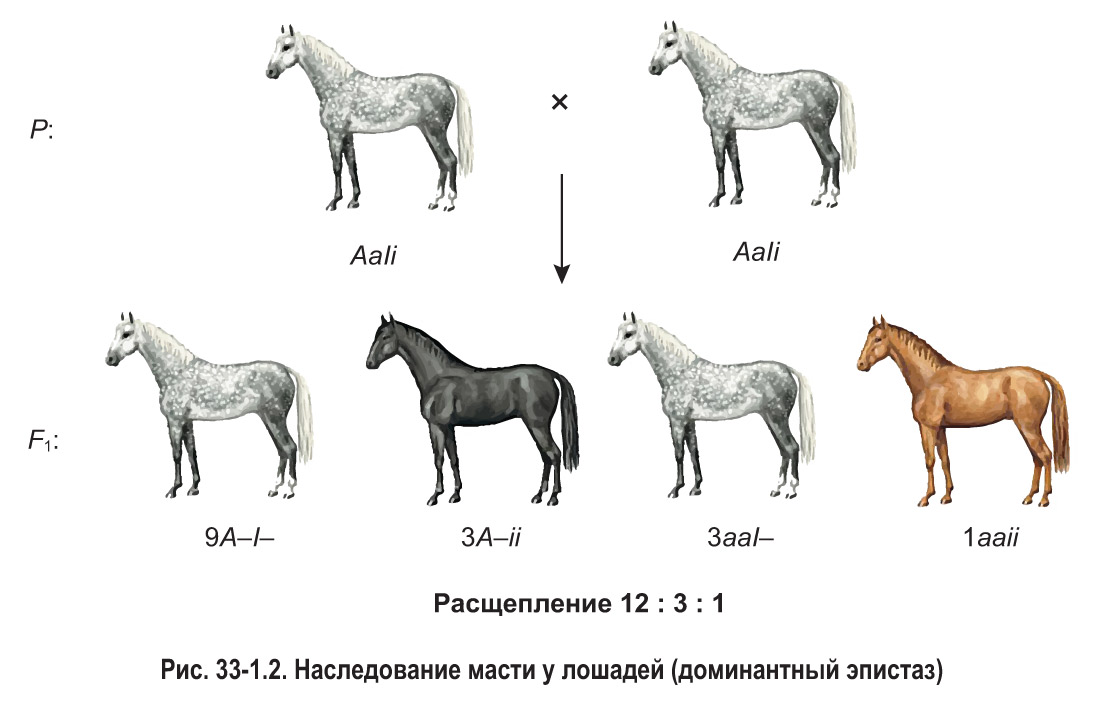
Примером доминантного эпистаза может служить взаимодействие двух аллелей, определяющих масть (окраску) лошадей. Так, доминантный ген одной аллельной пары (А) обусловливает вороную (черную) масть лошади, а рецессивный ген (а) — рыжую. Однако эти гены могут проявиться фенотипически только при отсутствии в генотипе лошади доминантного гена другой аллели — ингибитора (I). При наличии ингибитора проявление генов вороной и рыжей масти подавляется. В рассматриваемом примере ген-супрессор имеет собственное фенотипическое проявление: в случае его присутствия у особей развивается серая окраска.
Рассмотрим скрещивание дигетерозиготных лошадей (рис. 33-1.2). Они имеют серую масть, что обусловлено наличием в генотипе доминантного гена-супрессора (I). В потомстве от скрещивания наблюдается расщепление 12 : 3 : 1. Все особи, содержащие в генотипе эпистатический ген (9A—I— и 3aaI—), наследуют серую масть. У жеребят, не имеющих гена I, проявляется вороная либо рыжая окраска в соотношении 3 вороные (3A–ii) к 1 рыжей (1aaii).
Если ген-супрессор является рецессивным, то взаимодействие неаллельных генов протекает по типу рецессивного эпистаза (криптомерии). Такое взаимодействие наблюдается, например, при наследовании цвета семян фасоли. У этого растения красная окраска семян (А) доминирует над желто-коричневой (а). Проявление рецессивного ингибитора (i) ведет к блокировке действия генов красной и желто-коричневой окраски, и семена оказываются белыми. Однако действие рецессивного эпистатического гена может проявиться только при отсутствии в генотипе соответствующего ему доминантного аллеля (I). В случае наличия такого гена действие супрессора подавляется. Так, скрещивание дигетерозиготных растений фасоли приводит к расщеплению среди потомков в соотношении: с красными, с желто-коричневыми и с белыми семенами .

Эпистаз наблюдается при наследовании многих признаков живых организмов. При различных формах эпистаза и разных особенностях проявления генов расщепление в потомстве от скрещивания дигетерозиготных особей может составлять 13 : 3, 12 : 3 : 1 или 9 : 3 : 4.
Полимерия — взаимодействие, при котором гены двух или более аллелей проявляются сходным образом, определяя развитие одного и того же признака. В случае полимерии неаллельные гены, контролирующие тот или иной признак (полимерные гены), принято обозначать одинаковыми буквами с цифровыми индексами. При этом гены каждой аллельной пары имеют одинаковый индекс. Например: А1, а1 — доминантный и рецессивный аллели первой пары генов, А2, а2 — второй пары и т. д. Известны две разновидности полимерии.
При некумулятивной полимерии доминантный признак проявляется в полной мере при наличии в генотипе хотя бы одного доминантного гена. Иными словами, количество доминантных генов не влияет на степень выраженности признака. Так, у пастушьей сумки форма плодов определяется двумя парами полимерных генов. При этом рецессивный признак — овальные плоды проявляется только у растений, имеющих генотип а1а1а2а2. Если в генотипе присутствует хотя бы один доминантный ген (неважно какой именно — А1 или А2), растение обладает доминантным признаком — плодами треугольной формы.
Скрещивание доминантной и рецессивной дигомозигот (рис. 33-1.3) приводит к появлению в первом поколении дигетерозиготных гибридов с треугольными плодами (А1а1А2а2). Во втором поколении наблюдается расщепление в соотношении 15 : 1.

В случае кумулятивной полимерии степень выраженности доминантного признака определяется количеством доминантных генов в генотипе. Чем больше таких генов, тем сильнее проявляется данный признак. Например, у пшеницы определенных сортов окраска зерен контролируется тремя парами неаллельных генов. Растения с генотипом а1а1а2а2а3а3 имеют белые зерна, а обладающие генотипом А1А1А2А2А3А3 — темно-красные. У растений с другими генотипами наблюдаются различные промежуточные варианты окраски, зависящие от числа доминантных генов. Так, наличие в генотипе только одного доминантного гена обусловливает бледно-розовый цвет зерен, двух — розовый, трех — светло-красный и т. д.
Установлено, что по типу кумулятивной полимерии наследуются многие количественные признаки организмов: рост человека, цвет его кожи (т. е. количество меланина), яйценоскость кур, молочность коров, содержание сахарозы в корнеплодах сахарной свеклы, белков в эндосперме семян злаков и др.

Многие признаки живых организмов определяются взаимодействием неаллельных генов. Основными типами такого взаимодействия являются комплементарность, эпистаз и полимерия. При комплементарном действии доминантные гены разных аллелей вместе обусловливают появление нового признака, не определявшегося ни одним из этих генов по отдельности. В случае эпистаза ген одной аллели (эпистатический ген) подавляет проявление генов другой аллельной пары. В зависимости от того, доминантным или рецессивным является эпистатический ген, различают доминантный и рецессивный эпистаз. Полимерия заключается в том, что гены двух или более аллелей проявляются сходно, определяя развитие одного и того же признака. При кумулятивной полимерии степень проявления доминантного признака зависит от числа доминантных генов, при некумулятивной наличие даже одного доминантного гена ведет к полному проявлению доминантного признака.
1. Укажите типы взаимодействия неаллельных генов:
Кодоминирование , эпистаз, полимерия, полное доминирование, неполное доминирование, комплементарность.
2. Что представляет собой комплементарное взаимодействие неаллельных генов? Приведите примеры.
3. В чем заключается действие эпистатических генов? Чем доминантный эпистаз отличается от рецессивного?
4. Что такое полимерия? В чем состоит различие между кумулятивной и некумулятивной полимерией?
5. Какие варианты расщепления могут наблюдаться в потомстве от скрещивания дигетерозигот в случае взаимодействия неаллельных генов по типу: а) комплементарности; б) эпистаза; в) некумулятивной полимерии?
6*. У хомяков коричневая окраска шерсти определяется одним доминантным геном, а абрикосовая — другим. Данные гены расположены в разных парах хромосом, их рецессивные аллели приводят к появлению белой окраски. Если же в генотипе хомяка присутствуют оба неаллельных доминантных гена, появляется новый признак — черная окраска. От скрещивания абрикосового самца с коричневой самкой родился детеныш с белой шерстью. Установите генотипы родительских особей. Какое потомство можно ожидать у них в дальнейшем?
7*. Окраска шерсти собак породы лабрадор контролируется двумя аллелями, локализованными в разных парах хромосом. Гены одной аллельной пары обусловливают развитие черной или коричневой окраски, при этом черная масть полностью доминирует над коричневой. В другой аллельной паре рецессивный ген обладает эпистатическим действием. Он подавляет проявление генов черной и коричневой окраски, обусловливая развитие шерсти светло-бежевого цвета. Дигетерозиготную особь скрестили с рецессивной дигомозиготной. Какой фенотип имеют скрещиваемые особи? Какова вероятность появления в их потомстве щенков, имеющих: а) черную; б) коричневую; в) светло-бежевую шерсть?
8*. Допустим, что у растений определенного вида длина лепестков определяется двумя парами полимерных генов, расположенными в негомологичных хромосомах. Особи с длиной лепестков 40 мм имеют полностью доминантный генотип, а с длиной 20 мм — полностью рецессивный. Растения с другими генотипами имеют лепестки длиной 35, 30 или 25 мм. Какова длина лепестков у дигетерозиготных растений? Какое расщепление в потомстве будет наблюдаться при скрещивании этих растений между собой?
Читайте также: