
Завязь цветка после оплодотворения превращается
Обновлено: 08.07.2024
Опылением называется, собственно, перенос пыльцы с тычинок на рыльца пестиков. Различают самоопыление (автогамию: греч. авто — сам) и перекрестное опыление (аллогамию: греч. аллос — другой). В первом случае пыльца опыляет рыльце того же цветка, во втором — других цветков того же самого (гейтоногамия: греч. гейтон — сосед) или иных (ксеногамия: греч. ксенос — чужой) экземпляров. Ксеногамия биологически выгоднее прежде всего потому, что при этом возрастают возможности рекомбинаций генетического материала и это способствует увеличению внутривидового разнообразия и дальнейшей приспособительной эволюции. Однако и самоопыление имеет важное значение — для стабилизации признаков вида, в селекции — при выведении чистых линий и т. п.
Гейтоногамия встречается часто у растений с многоцветковыми соцветиями (например, у сложноцветных — перекрестное опыление цветков в пределах одной корзинки). С генетической точки зрения этот вариант равноценен автогамии, так как не происходит рекомбинаций индивидуальных признаков.
Дихогамия. Цветки нередко имеют те или иные приспособления, препятствующие самоопылению. Очень распространена дихогамия. Этим термином обозначают неодновременное созревание пыльников и рыльца. Более раннее вскрывание пыльников, когда рыльце еще не созрело, называют протерандрией, более раннее созревание рыльца — протерогинией.
Протерандрия наблюдается у гвоздичных, гераниевых, мальвовых, сложноцветных, многих лилейных и др.; протерогиния — у многих крестоцветных, розовых, осоковых и др. Протерандрия распространена шире, чем протерогиния, видимо, вследствие того, что тычинки, расположенные кнаружи от плодолистиков, начинают развиваться раньше них.
Гетеростилия. У некоторых растений одни экземпляры имеют цветки с длинными столбиками, другие — с короткими, а иногда имеются еще и третьи — со столбиками промежуточной длины соответственно, и пыльники расположены у одних ниже, у других — выше, а у третьих — на уровне рыльца. Это явление называется гетеростилией или разностолбчатостью. Гетеростилия, возможно, служит защитой от гейтоногамии.
Гетеростилия у первоцвета.
Другой пример иллюстрирует крайнюю степень специализации, при которой растение опыляется единственным видом насекомого и полностью зависит от последнего.
У инжира (Ficus carica), как и у других видов рода Ficus, так называемые в обиходе плоды представляют в действительности крайне своеобразные соцветия-синконии, у которых ось трансформирована в шарообразное или грушевидное тело (рис. 277).
Схема опыления инжира. (1— синконии с мужскими и галловыми цветками (каприфиги); 2— синконии с мужскими недоразвитыми и женскими цветками (фиги); ГЦв — галловые цветки; МЦв—мужские цветки; МНЦв— мужские недоразвитые цветки; ЖЦв — женские цветки.
ОПЛОДОТВОРЕНИЕ.
Развитие пыльцевой трубки. Пылинка, попав на рыльце пестика, при отсутствии тормозящих факторов прорастает; при этом ее содержимое, одетое интиной, выпячивается через поры в экзине и образует пыльцевую трубку.
Между опылением и оплодотворением у разных растений проходит весьма различное время. Так, у некоторых дубов этот период достигает 12—14 месяцев, у ольхи и орешника—3—4 месяцев, у орхидей — обычно несколько недель. У большинства растений данный период не превышает 1—2 суток, а у портулака он длится всего 3—4 ч, у недотроги — менее 1 ч, а у кок-сагыза — 15—45 мин. Скорость роста пыльцевой трубки составляет 35 мм/ч. Повышение температуры, как правило, ускоряет рост. Так, у дурмана при 33°С скорость роста была в 3 раза выше, чем при 11°С.
Ядро клетки пыльцевой трубки и генеративное ядро (или образовавшиеся из него 2 спермия) находятся на растущем конце пыльцевой трубки. Достигнув завязи, пыльцевая трубка направляется к семязачатку и проникает в него чаще всего через микропиле. Это явление носит название порогами и. Оболочка зародышевого мешка растворяется, соприкасаясь с кончиком пыльцевой трубки. В зародышевом мешке пыльцевая трубка растет по направлению к яйцеклетке. Оболочка на кончике пыльцевой трубки разрывается, и оттуда выходят 2 спермия, из которых один сливается с яйцеклеткой, а другой — с вторичным ядром зародышевого мешка или с одним из центральных ядер. Происходит так называемое двойное оплодотворение — характерная особенность покрытосеменных, не встречающаяся у голосеменных.
Двойное оплодотворение открыл в 1898 г. русский ботаник С. Г. Навашин на двух растениях семейства лилейных—Lilium martagon и Fritil-laria tenella. Впоследствии из оплодотворенной яйцеклетки развивается зародыш, а из канальцевой клетки с оплодотворенным вторичным ядром — эндосперм. Эндосперм покрытосеменных оказывается, стало быть, триплоидным и в противоположность голосеменным возникает лишь после оплодотворения. В этом состоит одно из принципиальных различий между голосеменными и покрытосеменными.
По форме спермин различны. Часто они палочковидные и даже червеобразные. С. Г. Навашин, наблюдая подобную форму спермиев на своих объектах, предположил, что спермин обладают способностью к самостоятельному движению, несмотря на отсутствие жгутиков. Некоторые данные как будто бы подтверждают это предположение. В то время как спермин движутся в пыльцевой трубке всегда__вперед, некоторые плазматические тяжи передвигаются в обратном направлении. Это возможно лишь в случае самостоятельного движения спермиев. Значительный интерес представляет выяснение вопроса, входит ли в яйцеклетку совместно с ядром мужской гаметы также ее цитоплазма. Однако наблюдение за живым зародышевым мешком методически весьма затруднительно. Большинство ученых считают, что мужская цитоплазма к моменту оплодотворения разрушается. Следует подчеркнуть, что доказательства так называемой внехромосомной наследственности основываются на участии в оплодотворении лишь женской цитоплазмы.
Кроме обычного способа проникновения пыльцевой трубки через микропиле — порогамии, известны иные варианты. Проникновение через хйлазу называют халазогамией. Впервые она была обнаружена в 1894 г. Трейбом у своеобразного австралийского рода Casuarina, а вскоре после этого — С. Г. Навашиным у березы. При халазогамии пыльцевая трубка растет через ткани рыльца и столбика до семяножки, а затем опять-таки в ткани последней до уровня середины семязачатка и далее по ткани обоих покровов она поворачивает к верхушке нуцеллюса, минуя микропиле и полость между покровами (рисунок 2). Предположили, что у халазогамных покрытосеменных пыльцевая трубка, как и у голосеменных, не способна еще к росту через полость завязи, и, стало быть, береза, казуарина и др. относятся к наиболее примитивным цветковым.
Схемы порогамии (1) и халазогамии (2, 3). (ПТ — путь пыльцевой трубки; ЗМ — зародышевый мешок)
В настоящее время, как правило, придерживаются иной точки зрения, рассматривая халазогамию более с физиологической, чем с филогенетической, точки зрения. Следует учитывать, что халазогамия известна и у сравнительно эволюционно развитых растений, например у крестоцветных, манжеток и др. Кроме того, у некоторых видов имеет место как порогамия, так и халазогамия.
Формирование зародыша. Оплодотворенная яйцеклетка переходит в состояние покоя, весьма различное по времени и зависящее отчасти от внешних условий. У сложноцветных и злаков этот период наиболее короткий (несколько часов). Первое деление сопровождается заложением поперечной перегородки. Клетку, отделяющуюся в направлении середины зародышевого мешка, называют терминальной, другую — базальной. Дальнейшие деления у различных растений происходят по-разному. У крестоцветных, например, базальная клетка делится поперек, а терминальная — вдоль, что приводит далее к возникновению проэмбрия (предзародыша).
Первые стадии развития зародыша у пастушьей сумки (Capsella bursapastoris): (БК—базальная клетка; ТК — терминальная клетка; Г — гаусториальная клетка подвеска.)
Пыльца оказывается на липком рыльце пестика и удерживается на его поверхности благодаря своей шереховатой оболочке. Здесь пыльцевое зерно начинает прорастать и превращается в пыльцевую трубку , которая через столбик попадает в завязь.

Одна из клеток пыльцевого зерна делится с образованием двух мужских гамет (спермиев) . Затем пыльцевая трубка дорастает до пыльцевхода, спермии попадают в зародышевый мешок. Один из них сливается с яйцеклеткой и образуется зигота. Второй спермий попадает внутрь зародышевого мешка и сливается с ядром центральной клетки.
У покрытосеменных, цветковых растений уникальный процесс оплодотворения. Именно уникальный. Ни один другой живой организм не проходит через процесс, который биологи называют «Двойное Оплодотворение«.
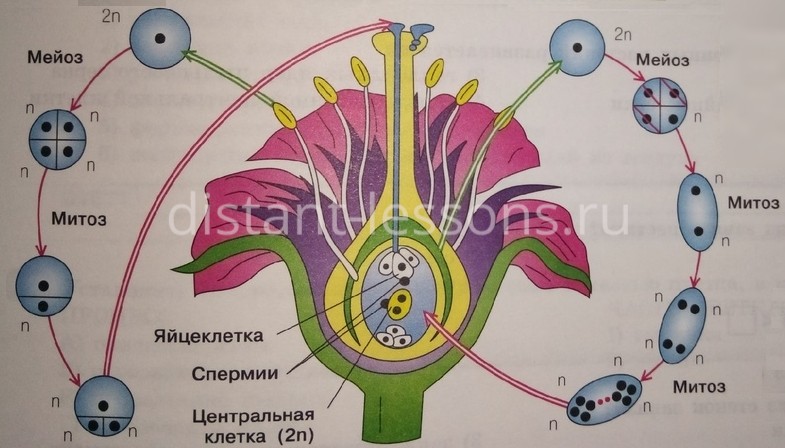
На женском гаметофите в семязачатке развивается мегаспора — женская половая клетка, которая формируется в завязи пестика.
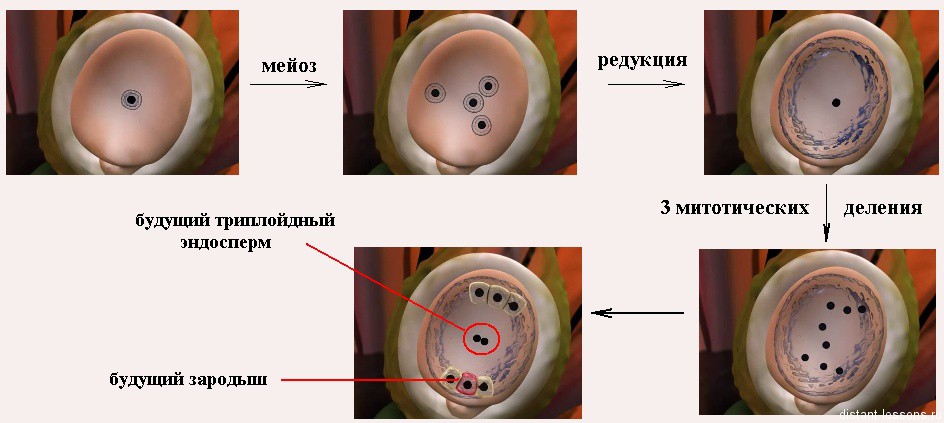
Спорогенная клетка (2n) сначала делится митозом. Образуются мегаспороциты (2n). Затем они делятся мейозом, образуются гаплоидные (1n) клетки — мегаспоры. У большинства видов 3 из этих клеток редуцируются и остается одна мегаспора.
Она развивается и ее ядро 3 раза делится митозом. Образуются 8 гаплоидных ядра, которые находятся в цитоплазме одной клетки. 6 ядер смещаются к краям семязачатка, 2 ядра остаются в центре клетки.
- Два слившихся ядра образуют центральное ядро,
- остающиеся на полюсах ядра превращаются в клетки:
— антиподы (1n),
— яйцеклетку (1n),
— синергиды (1n) - формируется женский гаметофит, или зародышевый мешок

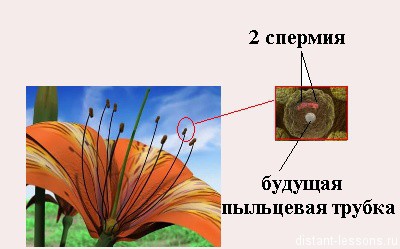
Мужские половые клетки — микроспоры.
В состав пыльцевого зерна входят 2 клетки:
- вегетативная клетка, которая в будущем формирует пыльцевую трубку и
- генеративная клетка, которая в будущем митозом образует 2 спермия.
Опыление — процесс переноса пыльцы с пыльников тычинок цветка на рыльце пестика
Пыльцевой зерно попадает на рыльце пестика, образующаяся пыльцевая трубка начинает прорастать внутрь столбика, стремясь к завязи. Когда пыльцевая трубка достигает зародышевого мешка, генеративная клетка образует 2 спермия. Один из спермиев оплодотворяет яйцеклетку, образуется зародыш. Второй спермий сливается с двумя ядрами в середине, образуя триплойдную клетку (3n) — эндосперм. Это и есть двойное оплодотворение.
Процесс двойного оплодотворения был открыт С.Г. Навашиным в 1898 г
Последовательность этапов этого процесса можно свести к схеме:
(такой вопрос часто встречается в части B ЕГЭ)

- после оплодотворения семязачаток превращается в семя
- завязь превращается в плод,
- из зиготы — зародыш семени
- из покровов семязачатка формируется семенная кожура.
Значение двойного оплодотворения
В семенах очень быстро (опережая развитие зародыша) образуются питательные ткани с запасом высокоэнергетических питательных веществ только после оплодотворения. У цветковых растений при развитии семязачатков не тратится время на создание питательных веществ, как у голосеменных растений, поэтому развиваются они гораздо быстрее. Таким образом, благодаря двойному оплодотворению ускоряется процесс формирования и семяпочки, и яйцеклетки, и семени
У голосеменных в оплодотворении участвует один спермий и эндосперм возникает до оплодотворения в результате деления мегаспоры, т.е. он гаплоидный и первичный.
Среди цветковых растений широко распространен апомиксис — образование зародыша без оплодотворения ( без участия мужской гаметы). У животных есть аналогичный процесс — партеногенез. Зародыш может развиваться из любых клеток зародышевого мешка: яйцеклетки, центрального ядра, синергид или антипод. Из остальных частей семязачатка образуются другие части семени.
Двудомные растения опыляются извне. Соответственно, это либо опыление животными (насекомыми, в основном), либо ветром.
Для привлечения насекомых растения используют яркую окраску цветка и аромат, пыльца крупная, липкая; ветроопыляемые растения обычно имеют невзрачные цветки, которые высоко поднимаются над землей и мелкую сухую пыльцу.
Перенос пыльцы из пыльника на рыльце пестика называется опылением. Различают два вида опыления: перекрестное и самоопыление.
При самоопылении рыльце принимает пыльцу того же цветка либо другого, но той же особи. Возможно опыление в закрытых, нераспустившихся цветках (горох). При перекрестном опылении переносится пыльца от разных особей. Это основной тип опыления цветковых растений (яблоня, ива, огурец и др.).

Перекрестное опыление
Перекрестное опыление осуществляется естественным (насекомыми, птицами, летучими мышами, ветром, водой) и искусственным (производит человек) путями.
Приспособленность растений к опылению ветром проявляется в наличии голых цветков, либо невзрачных, слабо развитых околоцветников. Они лишены нектарников и запаха, пыльцы образуют много, она легкая, сухая, мелкая, рыльца длинные, с большой поверхностью для улавливания пыльцы (рожь, кукуруза).
Приспособленность растений к опылению насекомыми характеризуется яркой окраской венчика, наличием нектарников, запаха (одуванчик, земляника). Пищей для насекомых являются нектар и пыльца. Окраска и запах служат для привлечения опылителей. Иногда цветки обладают запахом, характерным для самок насекомых того же вида. Это привлекает к ним самцов, которые и осуществляют опыление. Эволюция цветковых растений и их опылителей шла параллельно. Это так называемая сопряженная эволюция.

Приспособление растений к опылению насекомыми и ветром
Перекрестное опыление обеспечивает обмен генами, поддерживает высокую гетерозиготность популяций, дает материал для естественного отбора и сохраняет самое выносливое потомство — носителей наиболее благоприятного сочетания генов.
Искусственное опыление
Искусственное опыление производит человек для повышения урожая или получения новых сортов растений. При этом для нанесения пыльцы на рыльце пестика используют разные способы. Так, у кукурузы, имеющей однополые цветы, пыльцу собирают, стряхивая верхушечные метелки мужских цветков в бумажные воронки. Затем собранной пыльцой посыпают выступающие на верхушке початка длинные рыльца женских цветков.
При искусственном опылении подсолнечника стебли двух соседних растений наклоняют так, чтобы можно было прижать цветущую поверхность одной корзинки к другой. Можно переносить пыльцу, поочередно прижимая руку в варежке из мягкой материи к цветущим корзинкам разных растений.

Схема искусственного опыления
Для получения новых сортов растений с обоеполыми цветками необходима подготовка к искусственному опылению. Прежде всего из цветков растения, избранного в качестве материнского, еще в бутоне удаляют пыльники и защищают эти цветки марлевыми или бумажными мешочками от попадания пыльцы. Через 2-3 дня, когда бутоны раскроются, наносят на рыльца пестиков заготовленную пыльцу другого сорта чистой сухой акварельной кисточкой, мягким поролоном или кусочком резинки, прикрепленными к проволоке.
Двойное оплодотворение у цветковых растений
После опыления происходит процесс оплодотворения, но для этого нужен ряд условий: пыльца должна не только удержаться на рыльце, но и прорасти через столбик, достигнуть семязачатка и обеспечить слияние мужских клеток с женскими.
Двойное оплодотворение характерно для цветковых растений.
Обычно на рыльце попадает множество пыльцевых зерен. Они, как правило, имеют шероховатую поверхность и удерживаются липкой кожицей рыльца. Кроме этого, при попадании совместимой пыльцы клетки рыльца выделяют вещества, стимулирующие ее прорастание.

Схема двойного оплодотворения у цветковых растений
Начинается прорастание пыльцевых зерен с набухания. Затем через специальные поры (каналы) в наружной оболочке пыльцевого зерна внутренняя выпячивается в тонкую пыльцевую трубку, куда переходят вегетативное ядро и спермин. Пыльцевые трубки всех совместимых зерен, удержавшихся на рыльце пестика, растут по столбику, направляясь к семязачатку. Одна из них обгоняет в росте другие и, достигнув пыльцевхода, проникает через него к зародышевому мешку и здесь изливает в него свое содержимое.
Один из спермиев сливается с яйцеклеткой, а другой — со вторичным ядром центральной диплоидной клетки. Вегетативное ядро разрушается еще до проникновения пыльцевой трубки в зародышевый мешок.
Двойное оплодотворение у цветковых растений открыл русский цитолог и эмбриолог растений С.Г.Навашиным в 1898г.
При наличии в завязи семязачатков в каждом из них происходит вышеописанный процесс двойного оплодотворения. Называется он двойным потому, что сливаются две мужские клетки с двумя клетками женского гаметофита. В дальнейшем после оплодотворения в цветке начинается развитие семени и плода.
Образование семян
После оплодотворения внутри зародышевого мешка начинается быстрое митотическое деление триплоидного вторичного ядра, не имеющего периода покоя. Образуется большое количество ядер, затем между ними возникают, перегородки.
Эти вновь образовавшиеся клетки продолжают деление, заполняя всю полость зародышевого мешка питательной тканью — эндоспермом, который у одних растений полностью расходуется во время развития зародыша (бобовые, тыквенные), а у других — сохраняется в зрелых семенах (злаки). Одновременно происходит разрастание зародышевого мешка и семяпочки.
Формирование зародыша начинается с деления зиготы. После периода покоя зигота делится митотически на две клетки. Верхняя клетка, прилегающая к пыльцевходу, образует подвесок, отодвигающий нижнюю клетку в глубь эндосперма. Подвесок у одних видов растений остается одноклеточным, у других — делится поперечными перегородками и становится многоклеточным. Нижняя клетка разрастается в предзародыш семени сферической формы. Предзародыш делится на 4 клетки двумя перпендикулярными перегородками, затем каждая из этих клеток делится еще на две.
Сначала клетки более или менее однородны. По мере дальнейшего деления происходит дифференцировка клеток на зачаточный корешок, зачаточный стебель, зачаточные листочки (семядоли) и зачаточную почечку, окруженную семядолями. К этому времени семяпочка превращается в семя, ее покровы и остатки эндосперма образуют кожицу семени.
Таким образом, из оплодотворенной диплоидной яйцеклетки формируется зародыш семени, а из вторичной триплоидной клетки — питательная ткань — эндосперм, покровы семязачатка превращаются в покровы семени, а стенка завязи, разрастаясь, образует околоплодник.
Читайте также: