
Более раннее созревание и экспонирование рылец цветка до созревания тычинок называется
Обновлено: 07.07.2024
Сростные элементы формируются: 1) первоначально формируются отдельные бугорки, которые затем образуют сплошное кольцо; 2) при появлении элементы имеют широкое основание и срастаются в виде валика, позже делятся на зубчики, дольки.
Тычинки дифференцируются на нить и пыльник довольно поздно, пыльник формируется раньше нити. Нить удлиняется за счет интеркалярного роста. Число бугорков меньше числа тычинок, так как бугорки расщепляются. Плодолистики пестика похожи на листья, разрастаются внизу, срастаются, образуя завязь. Затем обособляется рыльце и столбик.
Цветение и опыление растений
Цветение – это период от распускания бугорков до засыхания околоцветника и тычинок. Функция цветения – обеспечение опыления и оплодотворения.
Периоды цветения различны. Существуют ремонтантныевиды, которые постоянно цветут и плодоносят (какао, лимон).
Растения, которые цветут многократно – поликарпические, те которые цветут 1 раз в жизни – монокарпические.
Интенсивность и срок первого цветения у разных растений различны. Некоторые цветут обильно периодически. У некоторых цветки зацветают: 1) раньше листьев; 2)одновременно с распусканием листьев. Цветки так же могут раскрываться в различное время суток.
Опыление – это процесс переноса пыльцы с пыльников тычинок на рыльце пестика.
Два типа опыления:
1. Самоопыления (или автогамия) – пыльца с тычинок попадает на рыльце пестика того же цветка.
2. Перекрестное (или аллогамия) – пыльца с тычинок одного цветка попадает на пестик другого цветка.
В 1750-51 гг. открыли перекрестное опыление Доббе и Мюллер, Большой вклад внесли в изучение опыления Й.Т. Кёльрёйтер и К.К. Шпренгель.
Самоопыление
Положительные стороны самоопыления:
ü обеспечивает образование семян и плодов без опылителя;
ü способствует стабилизации вида, сохраняет чистые линии
Есть клейстогамные цветки, которые не раскрываются (летние цветки фиалок, кислицы). Самоопыляющимися растениями являются – ячмень, пшеница, фасоль, овес, томат, лен.
Отрицательной стороной самоопыления является сужение приспособительной способности потомков.
Приспособления, исключающие самоопыление:
o самостерильность – неспособность пыльцы прорастать на рыльце пестика собственного цветка (рожь, рис, кукуруза и т.д.);
o дихогамия – неодновременное созревание в цветке пыльников и рылец: протерандрия – более раннее созревание тычинок, протерогиния – более раннее созревание рылец.
o гетеростилия (разностолбчатость) – различная длина пестиков и тычинок;
Перекрестное опыление
Виды перекрестного опыления:
a. гейтоногамия – пыльца с тычинок попадает на рыльце цветка в пределах соцветия или растения;
b. ксеногамия – попадает на растение одного вида;
c. гибридизация – попадает на цветок другого сорта или вида.
В зависимости от агентов опылителей: анемофилия (ветром), гидрофилия (водой), энтомофилия (насекомыми), орнитофилия (птицами), мирмикофилия (муравьями).
Положительной стороной перекрестного опыления является то, что в биологическом отношении оно более предпочтительное, так как приводит к комбинированию новых признаков. Отрицательной стороной – необходимость опылителей и благоприятных условий.
Энтомофилия – 80% растений опыляются насекомыми. Особенности растений, опыляемых насекомыми:
o пыльца с шипами, выростами, клейкая, зерна крупные, содержат около 30% белка;
o есть нектар – насекомым необходимо проникать внутрь цветка, а нектар выделяется в небольшом количестве, порциями; нектар и пыльца – первичные аттрактанты;
o ярко окрашенный венчик или венчиковидный околоцветник, мелкие цветки собраны в соцветия;
o запах – аромат или тухлый (индол); цвет и запах – вторичные аттрактанты.
Цветок первичных цветковых мало чем отличался от шишек голосеменных. Отличие лишь в том, что мегаспрофиллы цветковых превращены в плодолистики или карпеллы. Внутри плодолистика создаются новые условия для развития семезачатка и для процесса оплодотворения, возникает рыльцевая поверхность, что приводит к глубоким изменениям в развитии женского гаметофита.
Цветок возник из энтомофильного и обоеполого стробила, так как все наиболее примитивные цветы у тропических представителей цветковых опыляются насекомыми. В этом процессе ведущую роль сыграли насекомые, которые привлекались обилием пыльцы на шишках предков цветковых растений. Тем самым они способствовали перекрестному опылению. Одновременно они поедали семезачатки, что должно было привести к возникновению защиты семезачатков.
Внутри плодолистика семезачатки находят более надежную защиту.
Первыми опылителями цветковых были не специализированные лижущие и сосущие насекомые, лишенные хоботка. Многие считают, что это были жуки. Затем часть цветов опылялась ручейниками, наездниками и пилильщиками.
Образование цветочной трубки привело к возникновению бабочек и различных ос и пчел. Наиболее важной группой в процессе эволюции стали насекомые во всех возрастных состояниях, питающиеся пыльцой. Это привело к постоянству опыления и посещения цветков. Количество элементов цветка сократилось, и цветок приобрел специализированную форму. Возникла определенная изоляция отдельных популяций и в дальнейшем прооизошло образование новых видов. Огромное разнообразие цветковых растений (более 250 000 видов) в значительной степени обязано тем барьерам изоляции, которые возникли на основе энтомофилии. Исходными и примитивными считаются цветы с большим и неопределенным числом долей околоцветника, плодолистиков (пестиков), тычинок. Затем возникает некоторое определенное количество элементов в цветке и происходит срастание части элементов и на этой основе появляются зигоморфные цветы.
Опыление – необходимое условие для процесса оплодотворения, протекающего в цветке. Пыльца из пыльников так или иначе переносится на рыльце цветка. Различают два типа опыления – самоопыление и перекрестное опыление. Если пыльца переносится в пределах данного цветка или данной особи, то в этом случае происходит самоопыление. Различают разные формы самоопыления: автогамия, когда рыльце опыляется пыльцой того же цветка, гейтоногамию, когда рыльце опыляется пыльцой других цветков той же особи, и, наконец, клейстогамию, когда самоопыление происходит в закрытых, нераспускающихся цветках. Эти разные формы самоопыления в генетическом отношении вполне равноценны.
Если перенос пыльцы осуществляется между цветками разных особей, то в этом случае происходит перекрестное опыление. Перекрестное опыление – основной тип опыления цветковых растений. Он свойствен подавляющему большинству их.
В цветках весьма обычны специальные устройства морфологического характера, предотвращающие или по крайней мере ограничивающие самоопыление. Таковы двудомность, дихогамия, самонесовместимость, гетеростилия и др. Однако в них имеются также приспособления к самоопылению, способствующие последнему в том случае, когда перекрестное опыление по каким-либо причинам не произойдет. Иначе говоря, цветок допускает возможность не только перекрестного опыления, но и самоопыление.
Перекрестное опыление осуществляется следующими способами: с помощью насекомых (энтомофилия), летучих мышей (хироптерофилия), неживой природы – ветра (анемофилия) и воды (гидрофилия). В соответствии с этим можно говорить о биотическом и абиотическом опылении.
Перекрестное опыление обусловливает обмен генами и интеграцию мутаций, поддерживает высокий уровень гетерозиготности популяций, определяет единство и целостность вида. Это создает широкое поле для деятельности естественного отбора. Самоопыление, особенно постоянное, рассматривается как вторичное явление, вызванное крайними условиями окружающей среды, неблагоприятными для перекрестного опыления. Оно выполняет тогда страховую функцию. Постоянное самоопыление трактуется как тупик эволюционного развития. В этом случае происходит расщепление вида на серию чистых линий и затухание процессов микроэволюции. В этой правильной, но односторонней точке зрения на эволюционное значение самоопыления нашла отражение идея Дарвина, что природа питает отвращение к постоянному самооплодотворению. Это афоризм, как указывал сам Дарвин, будет ошибочным, если исключить из него слово “постоянному”. Указав на вредное действие постоянного самоопыления, Дарвин отнюдь не отрицал его значения вообще. По современным представлениям, для прогрессивной эволюции необходимо как свободное скрещивание, так и некоторое ограничение его. Перекрестное опыление повышает уровень гетерозиготности в популяции, а самоопыление, наоборот, вызывает гомозиготизацию ее. Самоопыление влечет за собой в сущности изоляцию новых форм. В этом и заключается положительное значение для эволюции сочетания в ряду поколений самоопыления и перекрестного опыления.

Обоеполость и энтомофильность цветка представляют первичное явление. В цветках первичных покрытосеменных наряду с весьма примитивной энтомофилией, вероятно, осуществлялось самоопыление. Обоеполость цветка способствовала самоопылению, поскольку приспособления к ограничению его ещё не были развиты. Разделение полов в цветке ограничивает или вполне исключает самоопыление. Оно привело к образованию разных половых типов цветковых растений.
Разделение полов как приспособление
К перекрестному опылению
Двудомность, по современным представлениям, у цветковых растений вторична. Это надежное средство предотвращения самоопыления. Эффективность такого механизма оплачивается дорогой ценой, поскольку половина популяции двудомных растений не дает семян. Примером могут служить ивы, тополя, осины, облепиха, спаржа, крапива двудомная и др.
Однодомность (женские и мужские цветки находятся на одной особи) встречается как у насекомоопыляемых, так и у ветроопыляемых растений. Примером однодомных растений могут служить представители тыквенных, кукуруза, большинство осок и др. Однодомность устраняет автогамию, но не предохраняет от равноценной ей гейтоногамии.
Иногда при внешней морфологической обоеполости цветки функционируют как однополые. В качестве примера могут служить кажущиеся обоеполыми цветки степного растения песчанки длиннолистной. У одних особей в цветках наблюдается редукция тычинок, и тогда они выступают как женские, у других отмечается дегенерация семязачатков. Такие цветки занимают как бы промежуточное положение между истинно обоеполыми и однополыми цветками.
Другие способы ограничения и предотвращения самоопыления.
Дихогамия – это функциональная раздельнополость. Она вызвана разновременным созреванием и экспонированием пыльцы и рыльца в цветках, вследствие чего последние выступают то в мужской, то в женской фазе. Дихогамия проявляется в форме протандрии (при более раннем созревании пыльцы) или протогинии (при более раннем созревании рылец). Одновременное созревание пыльцы и рылец в цветке носит название гомогамии. Дихогамия обычно рассматривается как приспособление к перекрестному опылению и как средство предотвращения самоопыления. Это традиционное представление о дихогамии нуждается в уточнении. Дихогамия необязательно исключает самоопыление. Поэтому значение дихогамии следует усматривать в том, что она более или менее ограничивает самоопыление, способствуя тем самым перекрестному опылению. Такое сочетание перекрестного опыления и самоопыления играет, как уже говорилось, положительную роль в эволюции.
Явление гетеростилии состоит в том, что у вида имеются две или три формы цветков, находящиеся на разных особях и различающиеся по длине столбиков и расположению тычинок (диморфные и триморфные). У диморфных первоцветов длинностолбчатая форма имеет цветки с длинным столбиком, рыльце которого находится в зеве венчика, а тычинки – ниже, в глубине венчика. У короткостолбчатой формы обратное расположение столбиков и пыльников. Формы гетеростильных растений различаются по другим признакам. Так, короткостолбчатые цветки характеризуются по сравнению с длинностолбчатыми более крупной пыльцой. Гетероморфная несовместимость встречается редко. Она известна у 56 родов из 23 семейств.
Насекомые сыграли выдающуюся роль не только в эволюции цветка, но и в его происхождении. Первичные покрытосеменные, очевидно, имели обоеполые энтомофильные цветки. Обоеполость цветка явилась предпосылкой для автогамии. Раздельнополость и способность к ветроопылению у покрытосеменных имеют вторичное происхождение. Первичные цветковые растения, видимо, не были узкоспециализированы по способу опыления, и энтомофилия у них была довольно примитивна. Вполне возможно, что наряду с насекомоопылением некоторую роль играло и ветроопыление или даже самоопыление. Примитивные цветковые растения ещё не обладают способностью к выделению нектара, и приманкой для насекомых служит исключительно пыльца. Перелом в развитии энтомофилии был связан с появлением нектарников и с выделением нектара в цветках. Нектар явился дополнительным к пыльце пищевым ресурсом, привлекавшим насекомых к цветку. Появление нектара вызвало коренное изменение состава опылителей. Если первоначальными опылителями были, вероятно, только жуки, то теперь большую роль в опылении стали играть высшие перепончатокрылые, двукрылые и чешуекрылые. Этот перелом в развитии энтомофилии подготовил быстрое возвышение цветковых растений в середине мелового периода.
Хорошо известно, что насекомые посещают цветки для сбора пищи (пыльцы и нектара). Но иногда играют роль другие причины – поиски убежища, отложение яиц в цветке, сбор строительного материала.
Пыльца является важнейшим источником пищи, которую насекомые находят в цветке. Она содержит белки, жиры, углеводы, зольные элементы, ферменты и витамины. Поэтому насекомые собирают пыльцу не только энтомофильных, но и анемофильных растений (орешник, злаки, подорожники), однако при этом они не принимают участия в их опылении. Следует, однако, заметить, что в качестве пищи пыльца энтомофильных растений представляет определенные преимущества – она более богата жирами и белками.
Цветки, привлекающие насекомых пыльцой, образуют огромное количество пыльцевых зерен (ветреница, мак, шиповник, зверобой). Мак и пион по размеру пыльцевой продукции превосходят многие анемофильные растения. У многих растений существует определенный дневной ритм в выделении пыльцы. Вскрытие пыльников и экспонирование пыльцы происходят либо в течение всего дня, либо в определенные сроки (утром, после полудня, в первую половину дня).
У энтомофильных растений пыльца относительно долговечна, но вода губительна для пыльцы. Известны разнообразные средства защиты ее от дождя и росы. Из них следует упомянуть ориентацию колокольчатых, бокальчатых и чашевидных цветков открытым зевом венчиком вниз. Такая ориентация у некоторых растений бывает постоянной и сохраняется на все время, пока пыльца нуждается в защите (черника, брусника, ландыш, спаржа, колокольчики), у других растений изгибание цветоножек и поникание цветков совершается периодически, перед дождем, в пасмурную погоду или ночью (адонис весенний, ветреница и др.). Такую же функцию защиты пыльцы играет закрывание цветков перед дождем, на ночь и в пасмурную погоду у горечавок, некоторых колокольчиков, кувшинок.
Нектар является важнейшим источником пищи для всех посетителей цветков. Он выделяется особыми экскреторными железами – нектарниками. Нектар представляет собой раствор сахаров – фруктозы, глюкозы, сахарозы, мальтозы и др. Концентрация сахаров в нектаре зависит от вида растений и условий среды. Обычно содержание сахаров в нектаре колеблется в пределах 30–50%, причем оно меняется в течение дня. Жидкий, а также высококонцентрированный нектар пчелы избегают, тогда как птицы–опылители в тропиках предпочитают водянистый нектар.
Форма и расположение цветковых нектарников весьма разнообразны. Они возникли независимо в разных филетических линиях покрытосеменных. Нектарники встречаются на разных органах цветка – чашелистиках, лепестках, цветоложе, тычиночных нитях, на наружной поверхности плодолистиков. Хорошо известны нектарные ямки у основания лепестков лютика. В цветках с нижней завязью они имеют вид диска, окружающего столбик у зонтичных, колокольчиковых, сложноцветковых.
Для привлечения насекомых служат, помимо пищи, также окраска и запах цветка, форма его имеет лишь небольшое значение. Заметность цветков определяется их окраской и цветовым контрастом с окружающей зеленью. Окраска цветков представляет полную цветовую гамму от длинноволновых лучей красного цвета до коротковолновых – ультрафиолетовых лучей, не воспринимаемых человеческим глазом, но воспринимаемых пчелами. Она обусловлена различными пигментами – флавоноидами, каротиноидами, антофеином, бетацианинами. Антоцианы растворены в клеточном соке, но иногда могут выпадать в клетках в форме кристаллов. Большинство окрасок цветов можно приписать им. В зависимости от реакции клеточного сока антоцианы вызывают красную окраску, синюю, фиолетовую. Желтая окраска цветов зависит от присутствия флавонов, их производных и каротиноидов. Последние определяют, например, желтую окраску цветков лютиков, купальницы, калужницы, чистотела, донника, зверобоя, подсолнечника и др. Коричневый пигмент антофеин вызывает черные пятна на лепестках. Белая окраска цветов не связана с наличием какого-либо особого пигмента, а обусловлена очень рыхлым строением лепестков вследствие наличия в них множества воздухоносных полостей. Кроме огромного разнообразия окрасок, доступных восприятию человека, цветки имеют ещё одну так называемую окраску, которая не воспринимается нами, но хорошо различается пчелами. Речь идет об ультрафиолетовом излучении цветков. Сильное ультрафиолетовое отражение обнаружено у цветков многих растений – дербенника иволистного, переступня и др. Очень невзрачные для нас цветки переступня хорошо различаются и интенсивно посещаются пчелами благодаря отражению ультрафиолетовых лучей.
У некоторых цветков наблюдается изменение окраски в течение жизни. Наиболее известным примером в этом отношении является медуница, у которой цветки при распускании бывают розово-пурпуровыми, а потом становятся синими. У чины весенней цветки сначала красные, а затем зеленовато-синие. Иногда меняется окраска какой-либо части цветка, как, например, у конского каштана, у которого желтое пятно на лепестках становится сначала оранжевым, а потом темно-красным, причем нектар выделяется только в начальной фазе.
Разнообразная окраска цветов, безусловно, имеет приспособительное значение, привлекая насекомых опылителей. Насекомые обладают способностью различать цвета, но их цветовое зрение, особенно у пчел, отличается рядом особенностей по сравнению с цветовым зрением человека. Главная особенность заключается в их восприимчивости к коротковолновым ультрафиолетовым лучам.
Пчелы отличаются слепотой к красной окраске, поэтому окраска цветов тюльпана, фасоли и мака они воспринимают как ультрафиолетовую.
Дневные бабочки обладают цветовым зрением. Цветки, опыляемые ночными бабочками, имеют обычно светлую окраску – белую, бледно-желтую и др. Цветки, опыляемые такими бабочками, источают к вечеру сильный аромат. Это дало повод думать, что запах является для ночных бабочек единственным средством ориентации.
Двукрылые посещают цветки ради пыльцы и нектара. В качестве посетителей и опылителей наиболее важны цветочные мухи. Посещаемые ими цветки имеют красную окраску.
В географическом распределении окраски цветов обнаруживаются некоторые закономерности. Обращает внимание обилие в тропиках ярких цветов, которые почти отсутствуют в средних широтах. Это связано с тем, что в тропиках большую роль в опылении играют птицы. В высокогорьях и приполярных областях растения с белыми цветками составляют до 40%, тогда как в Средней Европе таких растений около 23%, а в пустынях и сухих областях их ещё меньше.
Цветки привлекают насекомых–опылителей не только окраской, но часто также запахом. Источником запаха обычно являются лепестки, но также и тычинки, стаминодии и нектарники. В испускании запаха, как и в выделении нектарника, наблюдается суточная периодичность. Наиболее известным примером являются мыльнянка, смолевки, вечерница, лилии. У многих из этих растений цветки распускаются к вечеру и открыты ночью.
Эти факторы действуют совместно, но относительное их значение неодинаково на дальнем и близком расстоянии. Окраска цветка является в большинстве случаев фактором дальнего действия. Запах к цветку как фактор дальнего действия, за некоторыми исключениями, не играет существенной роли в прилете насекомых к цветку. Наоборот, он выступает как решающий химический фактор близкого действия.
Опылением называется, собственно, перенос пыльцы с тычинок на рыльца пестиков. Различают самоопыление (автогамию: греч. авто — сам) и перекрестное опыление (аллогамию: греч. аллос — другой). В первом случае пыльца опыляет рыльце того же цветка, во втором — других цветков того же самого (гейтоногамия: греч. гейтон — сосед) или иных (ксеногамия: греч. ксенос — чужой) экземпляров. Ксеногамия биологически выгоднее прежде всего потому, что при этом возрастают возможности рекомбинаций генетического материала и это способствует увеличению внутривидового разнообразия и дальнейшей приспособительной эволюции. Однако и самоопыление имеет важное значение — для стабилизации признаков вида, в селекции — при выведении чистых линий и т. п.
Гейтоногамия встречается часто у растений с многоцветковыми соцветиями (например, у сложноцветных — перекрестное опыление цветков в пределах одной корзинки). С генетической точки зрения этот вариант равноценен автогамии, так как не происходит рекомбинаций индивидуальных признаков.
Дихогамия. Цветки нередко имеют те или иные приспособления, препятствующие самоопылению. Очень распространена дихогамия. Этим термином обозначают неодновременное созревание пыльников и рыльца. Более раннее вскрывание пыльников, когда рыльце еще не созрело, называют протерандрией, более раннее созревание рыльца — протерогинией.
Протерандрия наблюдается у гвоздичных, гераниевых, мальвовых, сложноцветных, многих лилейных и др.; протерогиния — у многих крестоцветных, розовых, осоковых и др. Протерандрия распространена шире, чем протерогиния, видимо, вследствие того, что тычинки, расположенные кнаружи от плодолистиков, начинают развиваться раньше них.
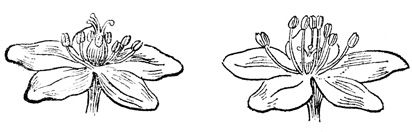
Гетеростилия. У некоторых растений одни экземпляры имеют цветки с длинными столбиками, другие — с короткими, а иногда имеются еще и третьи — со столбиками промежуточной длины соответственно, и пыльники расположены у одних ниже, у других — выше, а у третьих — на уровне рыльца. Это явление называется гетеростилией или разностолбчатостью. Гетеростилия, возможно, служит защитой от гейтоногамии.
Гетеростилия у первоцвета.
Другой пример иллюстрирует крайнюю степень специализации, при которой растение опыляется единственным видом насекомого и полностью зависит от последнего.
У инжира (Ficus carica), как и у других видов рода Ficus, так называемые в обиходе плоды представляют в действительности крайне своеобразные соцветия-синконии, у которых ось трансформирована в шарообразное или грушевидное тело (рис. 277).
Схема опыления инжира. (1— синконии с мужскими и галловыми цветками (каприфиги); 2— синконии с мужскими недоразвитыми и женскими цветками (фиги); ГЦв — галловые цветки; МЦв—мужские цветки; МНЦв— мужские недоразвитые цветки; ЖЦв — женские цветки.
ОПЛОДОТВОРЕНИЕ.
Развитие пыльцевой трубки. Пылинка, попав на рыльце пестика, при отсутствии тормозящих факторов прорастает; при этом ее содержимое, одетое интиной, выпячивается через поры в экзине и образует пыльцевую трубку.
Между опылением и оплодотворением у разных растений проходит весьма различное время. Так, у некоторых дубов этот период достигает 12—14 месяцев, у ольхи и орешника—3—4 месяцев, у орхидей — обычно несколько недель. У большинства растений данный период не превышает 1—2 суток, а у портулака он длится всего 3—4 ч, у недотроги — менее 1 ч, а у кок-сагыза — 15—45 мин. Скорость роста пыльцевой трубки составляет 35 мм/ч. Повышение температуры, как правило, ускоряет рост. Так, у дурмана при 33°С скорость роста была в 3 раза выше, чем при 11°С.
Ядро клетки пыльцевой трубки и генеративное ядро (или образовавшиеся из него 2 спермия) находятся на растущем конце пыльцевой трубки. Достигнув завязи, пыльцевая трубка направляется к семязачатку и проникает в него чаще всего через микропиле. Это явление носит название порогами и. Оболочка зародышевого мешка растворяется, соприкасаясь с кончиком пыльцевой трубки. В зародышевом мешке пыльцевая трубка растет по направлению к яйцеклетке. Оболочка на кончике пыльцевой трубки разрывается, и оттуда выходят 2 спермия, из которых один сливается с яйцеклеткой, а другой — с вторичным ядром зародышевого мешка или с одним из центральных ядер. Происходит так называемое двойное оплодотворение — характерная особенность покрытосеменных, не встречающаяся у голосеменных.
Двойное оплодотворение открыл в 1898 г. русский ботаник С. Г. Навашин на двух растениях семейства лилейных—Lilium martagon и Fritil-laria tenella. Впоследствии из оплодотворенной яйцеклетки развивается зародыш, а из канальцевой клетки с оплодотворенным вторичным ядром — эндосперм. Эндосперм покрытосеменных оказывается, стало быть, триплоидным и в противоположность голосеменным возникает лишь после оплодотворения. В этом состоит одно из принципиальных различий между голосеменными и покрытосеменными.
По форме спермин различны. Часто они палочковидные и даже червеобразные. С. Г. Навашин, наблюдая подобную форму спермиев на своих объектах, предположил, что спермин обладают способностью к самостоятельному движению, несмотря на отсутствие жгутиков. Некоторые данные как будто бы подтверждают это предположение. В то время как спермин движутся в пыльцевой трубке всегда__вперед, некоторые плазматические тяжи передвигаются в обратном направлении. Это возможно лишь в случае самостоятельного движения спермиев. Значительный интерес представляет выяснение вопроса, входит ли в яйцеклетку совместно с ядром мужской гаметы также ее цитоплазма. Однако наблюдение за живым зародышевым мешком методически весьма затруднительно. Большинство ученых считают, что мужская цитоплазма к моменту оплодотворения разрушается. Следует подчеркнуть, что доказательства так называемой внехромосомной наследственности основываются на участии в оплодотворении лишь женской цитоплазмы.
Кроме обычного способа проникновения пыльцевой трубки через микропиле — порогамии, известны иные варианты. Проникновение через хйлазу называют халазогамией. Впервые она была обнаружена в 1894 г. Трейбом у своеобразного австралийского рода Casuarina, а вскоре после этого — С. Г. Навашиным у березы. При халазогамии пыльцевая трубка растет через ткани рыльца и столбика до семяножки, а затем опять-таки в ткани последней до уровня середины семязачатка и далее по ткани обоих покровов она поворачивает к верхушке нуцеллюса, минуя микропиле и полость между покровами (рисунок 2). Предположили, что у халазогамных покрытосеменных пыльцевая трубка, как и у голосеменных, не способна еще к росту через полость завязи, и, стало быть, береза, казуарина и др. относятся к наиболее примитивным цветковым.
Схемы порогамии (1) и халазогамии (2, 3). (ПТ — путь пыльцевой трубки; ЗМ — зародышевый мешок)
В настоящее время, как правило, придерживаются иной точки зрения, рассматривая халазогамию более с физиологической, чем с филогенетической, точки зрения. Следует учитывать, что халазогамия известна и у сравнительно эволюционно развитых растений, например у крестоцветных, манжеток и др. Кроме того, у некоторых видов имеет место как порогамия, так и халазогамия.
Формирование зародыша. Оплодотворенная яйцеклетка переходит в состояние покоя, весьма различное по времени и зависящее отчасти от внешних условий. У сложноцветных и злаков этот период наиболее короткий (несколько часов). Первое деление сопровождается заложением поперечной перегородки. Клетку, отделяющуюся в направлении середины зародышевого мешка, называют терминальной, другую — базальной. Дальнейшие деления у различных растений происходят по-разному. У крестоцветных, например, базальная клетка делится поперек, а терминальная — вдоль, что приводит далее к возникновению проэмбрия (предзародыша).
Первые стадии развития зародыша у пастушьей сумки (Capsella bursapastoris): (БК—базальная клетка; ТК — терминальная клетка; Г — гаусториальная клетка подвеска.)

26.07.2018
Роль опыления в развитии и жизнедеятельности сельскохозяйственных культур огромна. Этот этап необходим растениям для образования семян и плодоношения, поэтому именно качественное опыление является решающим фактором в получении хороших урожаев. В процессе эволюции образовались две формы растений, отличающихся между собой типом опыления. Меньшая их часть относится к самоопыляемым, у которых процесс перенесения пыльцы с пыльников на рыльце происходит на одном и том же цветке. Большинство же культур (до 90%) входит в группу перекрёстноопыляемых растений, для которых характерно перенесение пыльцы с пыльников одного цветка на рыльце другого. В этом случае возможны два варианта: если оба цветка находятся на одном и том же растении, то процесс их опыления определяется как гейтоногамия, в сущности являющаяся одной из форм самоопыления; если опыляются цветки, расположенные на разных растениях, то имеет место ксеногамия.
Перекрестное опыление появилось в результате эволюционного развития как более прогрессивное, при котором образуется значительно большее количество генетических комбинаций, позволяющих получить разнообразные генотипы следующих поколений, что значительно увеличивает уровень приспосабливаемости растений к условиям существования и повышает их жизнеспособность. Самоопыление ведёт к получению малопрогрессивного потомства, склонного к вырождению, но в селекционной работе этот тип опыления имеет большое значение, так как помогает сохранить чистоту вида.


Дихогамия подразумевает разновременность начала и окончания функционирования тычинок и пестиков: более раннее созревание рыльца, чем пыльников (протерогиния) или обратную ситуацию, когда вскрытие пыльников опережает созревание рыльца (протерандрия). Протерогиния встречается у многих ветроопыляемых растений (злаки, полыни, подорожниковые, осоки), как правило, представителей семейств Крестоцветных (лат. Crucíferae (Brassicaceae)), Розовых (лат. Rosáceae), Барбарисовых (лат. Berberidáceae), Осоковых (лат. Cyperáceae). Более распространенная в природе протерандрия характерна для растений семейств Зонтичных (лат. Umbellíferae), Сложноцветных (лат. Compósitae), Гвоздичных (лат. Caryophylláceae), Колокольчиковых (лат. Campanuláceae), Мальвовых (лат. Malvaceae (Bombacaceae)), Лилейных (лат. Liliaceae), Гераниевых (лат. Geraniáceae).

Гетеростилия (разностолбчатость) проявляется у растений одного вида как наличие цветков с неодинаковой длины столбиками пестиков и тычиночными нитями, в результате чего рыльца и пыльники располагаются на разной высоте, что препятствует самоопылению цветка. Гетеростилию с диморфными цветами можно наблюдать у медуницы, примулы, гречихи. Половина растений этих видов имеет цветы с высокими столбиками, а половина – с низкими. Встречаются также виды растений (кислица, дербенник иволистный) с триморфными цветами, когда часть их экземпляров имеет высокие столбики, часть – низкие, а часть – столбики промежуточной высоты.
Культуры перекрёстноопыляемые, находящиеся в зависимости от насекомых-опылителей, обладают некоторыми особенностями для привлечения пчел, шмелей и других полезных насекомых. Часть таких растений выделяют сильные ароматические вещества, некоторые из них отличаются яркой окраской лепестков или других частей, у других представителей наблюдается формирование крупных, привлекающих внимание соцветий или цветов необычной формы. Но самым важным фактором, привлекающим насекомых, является наличие в цветках необходимого им пищевого продукта – нектара и пыльцы.

В то же время эта особенность строения цветков ставит культуру в полную зависимость от участия в процессе опыления насекомых-опылителей, а именно пчел. Практика показывает, что 70% опыления цветов гречихи обеспечиваются пчелами, и лишь 30% - другими насекомыми. Разная высота расположения пыльников и рылец способствует лучшему соприкосновению пчелы с пыльцой во время сбора нектара. Перемещаясь на цветок с иным расположением пыльников, насекомые оставляют часть пыльцы на рыльце, осуществляя таким образом перекрестное опыление растения.

Посевы из семян первого и второго поколения, полученных в результате опыления пчелами, отличаются лучшим ростом и развитием. Чтобы обеспечить получение высоких урожаев, к началу цветения гречихи необходимо подвозить на поля пчелиные пасеки из расчета 2 семьи на 1 га посевов, что позволяет провести насыщенное опыление растений. На больших площадях практикуют встречное опыление.
Читайте также: